От хаоса наблюдений к динамической типологии: обсуждение на примере популяционных систем зелёных лягушек Дмитрий Шабанов
От хаоса наблюдений к динамической типологии: обсуждение на примере популяционных систем зелёных лягушек
Дмитрий Шабанов
Опубликовано 11 июля 2013
Выложив очередную колонку, я раз за разом заглядываю на сайт КТ: как восприняли? Чаще сначала появляются комментарии от тех, кто понял, а потом подтягиваются пропагандисты креационизма и сакральных традиций, а также профессиональные спорщики. Реагировать? Надо понять, для кого я пишу колонки. Естественно, для читателей (ну и, конечно, немного для себя). Значит, надо отвечать. Как это делать, не утопая в грызне под колонками? Тут я попробую рассказать об одной работе с моим участием. Сможет ли она стать аргументом, позволяющим проиллюстрировать подход, который кажется мне правильным?
Я расскажу о фрагменте работы, которую три недели назад защитила как кандидатскую Марина Кравченко. Я — научный руководитель этой работы. Главные (с моей точки зрения) её результаты опубликованы с моим участием (здесь — сама модель, а тут — результаты моделирования, о которых я расскажу).
То, что я сейчас буду описывать, не является сутью диссертации Марины с точки зрения формальных требований. Зато это та её часть, из которой, как мне кажется, следуют определенные мировоззренческие выводы.
Марина Кравченко изучала трансформации популяционных зелёных лягушек. Динамика этих систем (мы называем их ГПС, гемиклональными популяционными системами) практически не изучена. Почему? Сейчас объясню.
Начну с утверждения, что основные свойства хорошо известных нам биосистем (организмов, популяций, сообществ) являются следствием передачи генетической информации с рекомбинацией (перебором сочетаний).
Главный механизм эволюции — отбор организмов, поколение за поколением проходящих непростой путь самоорганизации под влиянием наследственной программы. Одним из важных способов повышения эффективности эволюции таких наследственных программ стало оплодотворение — слияние клеток двух разных организмов. Каждая из этих клеток несет наследственные задатки, отобранные в предыдущих поколениях. Но у клетки, которая получится от их слияния, набор генетической информации окажется удвоенным. Поэтому неотъемлемой частью жизненных циклов с оплодотворением становится мейоз — клеточное деление, при котором количество генетической информации сокращается вдвое. Чередуясь в жизненном цикле, мейоз и оплодотворение поддерживают количество передаваемой генетической информации относительно постоянным.
Мейоз и оплодотворение поддерживают рекомбинацию генетического материала, ускоряя выработку приспособлений в эволюции. Генетически уникальные организмы, производящие генетически уникальные половые клетки, популяции с общим генофондом, межвидовая изоляция и особая эволюционная судьба каждого вида — следствия этого процесса.
Среднеевропейские зелёные лягушки (и несколько иных гибридогенных комплексов видов) вносят принципиальное изменение в базовый процесс. В этих комплексах образуются межвидовые гибриды, которые передают будущим поколениям комплекс генетической информации, полученной от одного из родительских видов, как единое целое, без рекомбинации. И в силу этого свойства всех иерархических структур, существование которых поддерживается размножением с рекомбинацией, меняются.
Организмы начинают производить одинаковые, клональные половые клетки. Гибриды принадлежат к полуклонам с функционально различающимися геномами (комплексами наследственной информации): одни из этих геномов передаются из поколения в поколение клонально, другие — используются в течение жизни особи, а потом бесследно уничтожаются. Границы между популяциями размываются, ведь в совместном размножении участвуют и представители родительских видов. Сплошь и рядом нарушение базового способа передачи генетической информации приводит к появлению необычных особей. Это полиплоиды с увеличенным количеством геномов, а также частичные межвидовые рекомбинанты. Между родительскими видами происходит обмен генетической информации, причем внедрение чужеродных генов затрагивает и те популяции, которые не участвуют в гибридизации.
Интереснейшая для популяционного эколога проблема — выяснение того, как поддерживается устойчивость популяционных систем, в рамках которых происходят такие чудеса. Как исследовать такую проблему?
Конечно, основа для такого ответа — сбор эмпирических данных. И нами, и другими исследователями установлено, что состав ГПС может со временем меняться. Некоторые из таких изменений мы с нашими студентами наблюдаем в поле. В любом случае такие трансформации — чрезвычайно длительный процесс, а их отслеживание — тяжёлая работа. О том, чтобы с помощью полевых исследований за разумное время изучить закономерности этих трансформаций, рассмотреть совокупность возможных состояний и логику их преобразования, говорить не приходится. Что же делать?
Применять моделирование. Причем не аналитическое, в ходе которого мы используем заранее установленные зависимости, описывающие результаты интересующих нас процессов, а имитационное. В ходе имитационного моделирования мы пытаемся хотя бы приблизительно описать этапы изучаемых преобразований, определяя, в каком направлении изменяется интересующая нас система.
Так мы и сделали. Описали годичный цикл зелёных лягушек, выделили в нём этапы, влияющие на состав ГПС, задали зависимости, характеризующие такие изменения. На основании собственных и литературных данных предположили, какой состав половых клеток характерен для каждой из форм гибридных лягушек. Получили конструкцию из уравнений, описывающих интересующие нас параметры.
Эта конструкция оказалась реализована в двух разных моделях. Мы с Мариной сделали и использовали модель на разностных уравнениях, написанную в Excel, а наши коллеги-математики собрали более мощную реализацию на основе профессиональных языков и пакетов. Профессиональная модель значительно превосходит нашу, тренировочную, по мощности, но заметно уступает ей по удобству пользования и перестройки. Марина защитила диссертацию, сделанную на тренировочной модели, но при этом мы с коллегами-математиками продолжаем работать над моделью «большой».
Расскажу об экспериментах с той моделью, которую назвал «тренировочной», Excel’евской. В ней прописан механизм размножения, роста и конкуренции лягушек с разными наборами геномов. Этот механизм носит вполне «общий» характер. Но кроме того, в модели есть блок, который обеспечивает возможность задавать, какие половые клетки производят представители того или иного генотипа. Тут-то и можно предусмотреть полуклональное наследование, характерное для зелёных лягушек.
А теперь, чтобы объяснить основную идею колонки, мне придется рассказать о нескольких экспериментах с моделью и объяснить логику её перестройки. Начну с примера, который подробно разбирал в колонке о Невидимой Ноге. В примерах, которые я привожу, буквы, L и R — обозначения геномов (каждый из 13 хромосом). Геномы, обозначения которых не берутся в скобки, — обычные, рекомбинантные, такие, как у каждого из нас (тот комплекс наследственной информации, что мы передаём детям, является относительно случайной смесью геномов, которые мы получили от отца и матери). В скобки берутся клональные геномы, которые передаются как единое целое.
Рассмотрим популяцию родительского вида, все скрещивания в которой таковы:
?RR ? ?RR ? ?RR : ?RR.
Здесь происходит примерно то же самое, что и в популяциях людей (и система определения пола та же): особи определенного вида производят особей того же вида. Но представим себе, что в такую популяцию попадает хотя бы одна гибридная особь, несущая клональный геном другого вида — например, ?(L)R. Заключением генома (L) в скобки мы указываем, что он попадает во все половые клетки. Раз так, всё потомство от скрещивания такого гибрида с особями родительского вида будет гибридным:
?(L)R ? ?RR ? ?(L)R : ?(L)R.
Теперь мы уже имеем дело не с популяцией, а с ГПС — гемиклональной популяционной системой, ведь в ней передаются из поколения в поколение клональные и рекомбинантные геномы, принадлежащие к разным видам. При скрещивании самца-гибрида с самкой родительских видов также возникнет только гибридное потомство (поскольку геном женский, всё потомство будет самками):
?RR ? ?(L)R ? ?(L)R.
Наконец, еще одной особенностью гибридов является то, что потомки от их скрещивания будут нежизнеспособными:
?(L)R ? ?(L)R ? ?(L)(L) ? †.
Итак, скрещивание «родителей» (особей родительского вида) порождает «родителей». От скрещивания «родителей» и гибридов возникают только гибриды, вследствие чего доля последних растет. Скрещивания гибридов не оказывают влияния на состав будущих поколений, так как потомство от них гибнет.
Результатом является то, что доля гибридов в такой ГПС возрастает, пока в ней не остаются только они. После этого такая система гибнет!
Многих людей удивляет, что для гибели родительской популяции достаточно попадания в неё одной-единственной гибридной особи. Тут надо думать не о вкладе в начальный состав представителей разных форм, а о том, присутствует ли в системе «инфекция», то есть геном, распространение которого ведёт к катастрофе.
Давайте посмотрим, какую траекторию опишет ГПС на фазовой плоскости (положение на которой отражает состав системы).
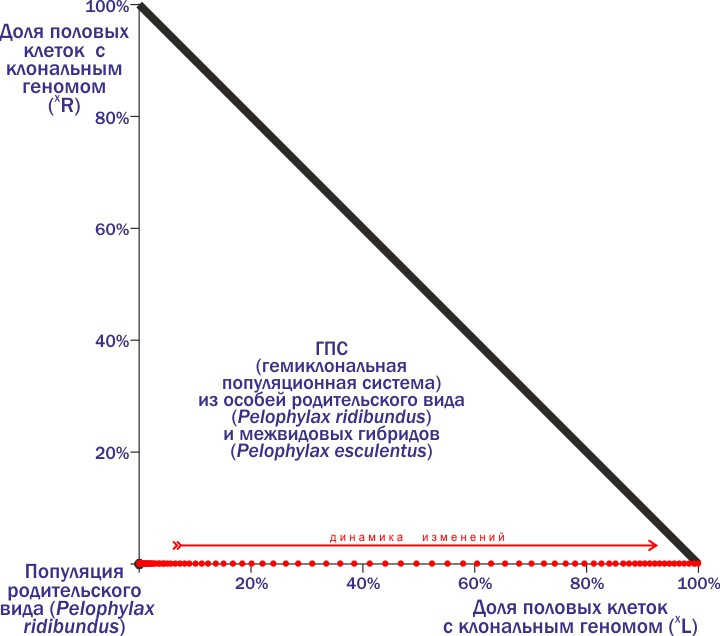
Смотрите. В углу — популяция родительского вида, с которой мы начали эксперимент. По осям — доли клональных геномов. В рассмотренном примере мы добавляем в популяцию из особей RR одну особь (L)R. В силу только что описанного нами механизма доля особей с геномом (L) в рассматриваемой ГПС начинает возрастать. Её траекторию маркируют красные точки; направление динамики показано стрелкой. Заканчивается всё гибелью ГПС при её попадании в правый угол: если 100% её состава — носители генома (L), все скрещивания приводят к появлению нежизнеспособного потомства.
При добавлении в популяцию родительского вида носителей геномов (R) ничего интересного не происходит. От скрещивания таких гибридов с особями родительского вида получаются «родители». Значит, популяция родительского вида находится в устойчивом равновесии относительно попадания в неё геномов (R) и в неустойчивом — относительно геномов (L).
А что будет, если геном (R) попадет в популяцию, где большинство особей являются носителями генома (L)?
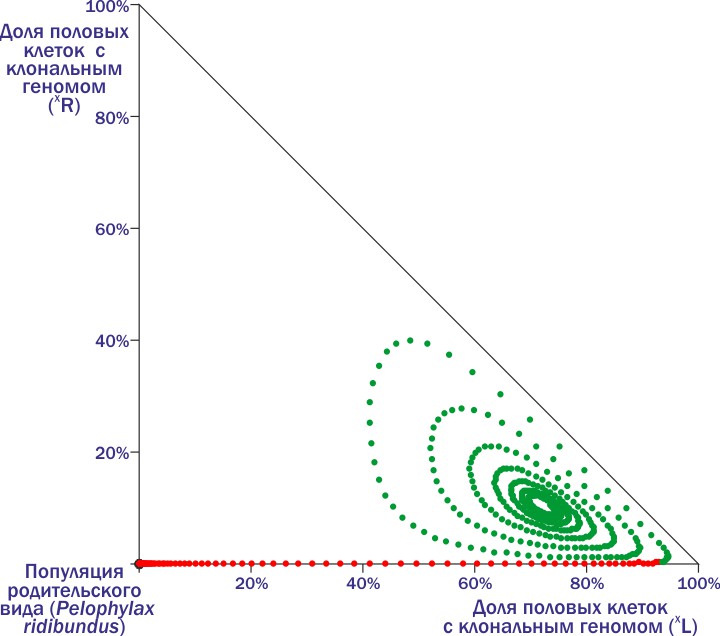
В том месте, где красные точки сменяются зелёными, в ГПС добавлена одна особь с клональным геномом (R). Описывая петли, модельная ГПС движется к состоянию устойчивого равновесия — где-то там, в центре, к которому стремится её траектория. И чтобы попасть туда, ГПС должна получить геном (R) во вполне определенный промежуток времени. Раньше — потеряется. Позже — хотя и вызовет резкие изменения состава, но все равно не спасёт систему от гибели.
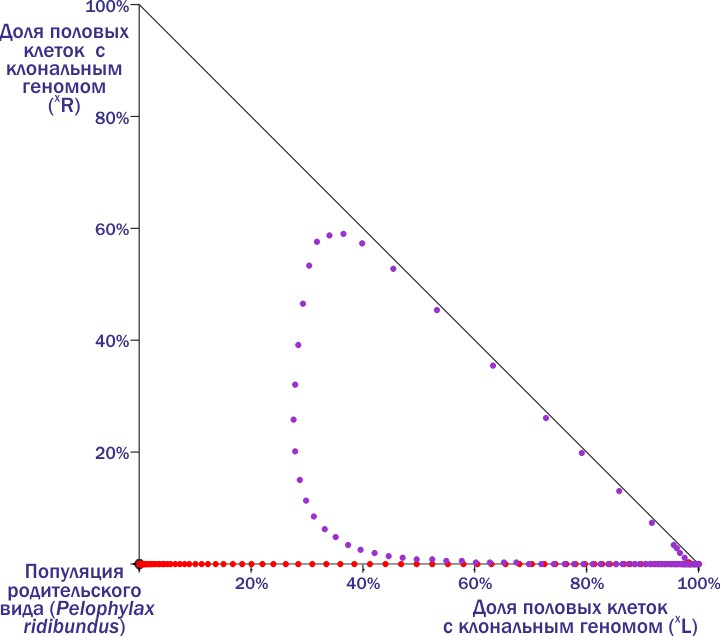
Вот тут в момент, когда красные точки сменяются фиолетовыми, в ГПС попадает носитель (R). Траектория системы выписывает петлю… и всё равно приходит её к гибели.
Сравнивая два предыдущих рисунка, мы можем увидеть, что снаружи от самой широкой зеленой петли на предпоследнем рисунке и внутри от фиолетовой петли на последнем из них проходит граница между областями, откуда ГПС попадает в два разных итоговых состояния — устойчивого равновесия и гибели. Этим состояниям соответствуют аттракторные (притягивающие к себе динамическую систему) точки на фазовой плоскости.
Раз так, мы можем разделить фазовую плоскость на бассейны разных аттракторных состояний!
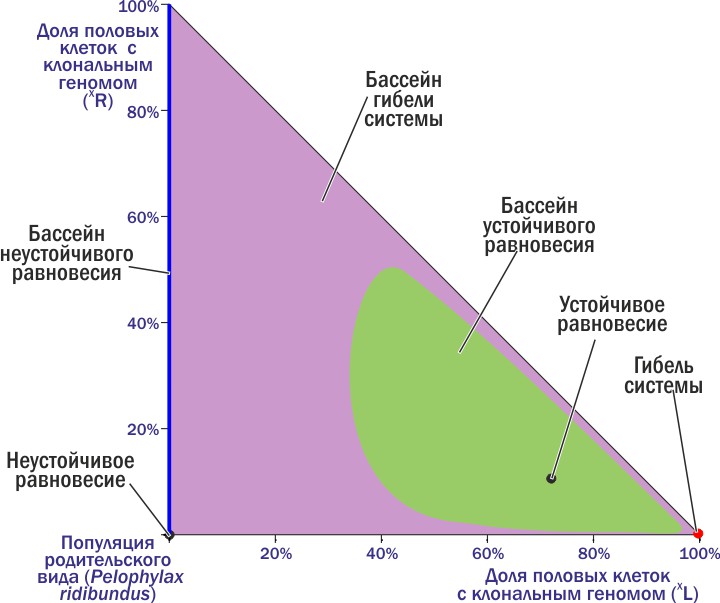
До сих пор различные ГПС классифицировали по их составу. Описанный здесь подход позволяет выделять динамические типы, которые характеризуются не только определенным составом, но и общностью ожидаемой судьбы. Эти типы — бассейны на фазовом пространстве состояний интересующих нас систем.
Смотрите, что мы сделали. Мы не вносили в обсуждение динамики интересующей нас системы никаких внешних критериев «хорошо — плохо». Мы просто установили, что одни состояния закономерно переходят в другие, и разделили их совокупность на группы. Теперь мы можем надеяться, что выделенные нами динамические типы отражают действительные особенности изучаемых систем. Мы можем выбрать, какие состояния являются для нас оптимальными. Дальше останется только понять, каким образом переходить от наблюдаемых состояний к желаемым (если такие появятся).
Спорщики доказывали мне в комментариях под предыдущими колонками, что даже здорового человека от больного нельзя отличить, не узнав, чего хочет Бог, а уж тем более нельзя понять, к какому возможному в будущем состоянию биосферы нам следует стремиться. Приведённый здесь пример показывает, как я надеюсь, что типология состояний сложной системы вполне может быть построена без ссылок на сверхъестественные целеуказания.
Да, мой пример касается далеко не только ГПС. Те типологии многих «будничных» динамических систем, которыми мы пользуемся, получены в ходе подобных по сути процедур. Разница только в том, что анализ пространства возможных состояний и путей перехода по нему чаще всего не требует имитационных моделей, реализованных на каком-то «железе», помимо нашего собственного мозга. Эти операции проводились на моделях, которые создает наша психика — частично осознанно, частично интуитивно. Вероятно, во многих случаях формализованные математические модели будут просто подтверждать результаты «мягкого», неформального моделирования. Однако есть ситуации, в которых наша способность к рациональному прогнозу или интуитивной оценке недостаточна. Что же, я надеюсь, что в таких случаях будет полезен описанный здесь подход.
Не «подпирайте» свои суждения ссылками на волю Творца. Давайте сами, силами нашего разума наблюдать, экспериментировать, моделировать, строить прогнозы и выбирать желаемое для нас состояние!
К оглавлению
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Специфичный механизм регуляции или отбор? Обсуждение одной гипотезы о механизмах воспроизводства межвидовых гибридов зелёных лягушек Дмитрий Шабанов
Специфичный механизм регуляции или отбор? Обсуждение одной гипотезы о механизмах воспроизводства межвидовых гибридов зелёных лягушек Дмитрий Шабанов Опубликовано 31 мая 2013 Несколько дней назад поспорил я со своим коллегой. Обсуждая одни и те же
Дмитрий Шабанов: Кофе и со‑знание ноосферы Дмитрий Шабанов
Дмитрий Шабанов: Кофе и со?знание ноосферы Дмитрий Шабанов Опубликовано 10 августа 2011 года Всё, пора прекращать «кофейные» колонки. Это – последняя.Мы увидели, какие источники пополняют энергию в чашке кофе и какие энергетические потоки организуют
Дмитрий Шабанов: О различиях между полами Дмитрий Шабанов
Дмитрий Шабанов: О различиях между полами Дмитрий Шабанов Опубликовано 11 мая 2011 года Недавно я сравнивал три основных типа размножения и кратко разбирался, почему воспроизводство с рекомбинацией благодаря половому процессу «лучше» (эволюционно
Дмитрий Шабанов: Фотопериодические реакции Дмитрий Шабанов
Дмитрий Шабанов: Фотопериодические реакции Дмитрий Шабанов Опубликовано 02 ноября 2011 года Мне уже приходилось писать, что меня огорчает недооценка нашей биологической природы. Биологический фундамент, на котором вырастает и наша культура, и
Дмитрий Шабанов: Блуждание глазами по всему небу Дмитрий Шабанов
Дмитрий Шабанов: Блуждание глазами по всему небу Дмитрий Шабанов Опубликовано 23 августа 2011 года Ибо весьма часто случается, что даже и нехристианин знает кое-что о земле, небе и остальных элементах видимого мира, о движении и обращении, даже
Дмитрий Шабанов: Почему есть мужчины и женщины? Дмитрий Шабанов
Дмитрий Шабанов: Почему есть мужчины и женщины? Дмитрий Шабанов Опубликовано 21 апреля 2011 года Я пообещал обсуждать биологические корни нашего поведения, а потом стал писать о другом — об учебниках, о "зелёных"... Исправляюсь.Мы так привыкли, что
Дмитрий Шабанов: Об извращённой любви к природе Дмитрий Шабанов
Дмитрий Шабанов: Об извращённой любви к природе Дмитрий Шабанов Опубликовано 18 мая 2011 года Я писал уже, что из-за активистов-природоохранников мне бывает стыдно называть себя экологом. Если вы составили представление об экологии по материалам
Дмитрий Шабанов: Энергия утреннего кофе Дмитрий Шабанов
Дмитрий Шабанов: Энергия утреннего кофе Дмитрий Шабанов Опубликовано 06 июля 2011 года Многие мои колонки посвящены биологической основе поведения людей. Увы, раз за разом я спотыкаюсь об одно непонимание. Многие читатели думают, что раз я говорю о
Дмитрий Шабанов: Что управляет нашим поведением? Дмитрий Шабанов
Дмитрий Шабанов: Что управляет нашим поведением? Дмитрий Шабанов Опубликовано 18 июля 2012 года Уже во многих своих колонках я сравнивал поведение человека с поведением других животных. Раз за разом сталкивался я с одной и той же реакцией: ну как же
Дмитрий Шабанов: Что отражается в чашке кофе? Дмитрий Шабанов
Дмитрий Шабанов: Что отражается в чашке кофе? Дмитрий Шабанов Опубликовано 12 июля 2011 года В прошлой колонке мы перечислили источники, пополняющие энергию утренней чашки кофе, как и энергию множества других процессов и результатов деятельности
Обсуждение перехода от гермафродитизма к раздельнополости как пример неклассического развития естественнонаучной гипотезы Дмитрий Шабанов
Обсуждение перехода от гермафродитизма к раздельнополости как пример неклассического развития естественнонаучной гипотезы Дмитрий Шабанов Опубликовано 02 февраля 2014 Я рад тому, как была принята позапрошлая колонка (не прошлая, политическая, а
Дмитрий Шабанов: Эволюция: биологическая и социальная Дмитрий Шабанов
Дмитрий Шабанов: Эволюция: биологическая и социальная Дмитрий Шабанов Опубликовано 08 ноября 2012 года Некоторым читателям может показаться странным фокус моего внимания в этой колонке. Я хочу разобраться в том, чем социальная эволюция отличается от
Дмитрий Шабанов: Судьба точек роста в системе декоративного образования Дмитрий Шабанов
Дмитрий Шабанов: Судьба точек роста в системе декоративного образования Дмитрий Шабанов Опубликовано 05 июля 2012 года Ф-ф-ф-у-у-у-х! Учебная практика на биостанции, где я провёл последний месяц, закончилась. Прощальный момент был таков: начальник
Дмитрий Шабанов: «Экологические задачи» для студентов и школьников Дмитрий Шабанов
Дмитрий Шабанов: «Экологические задачи» для студентов и школьников Дмитрий Шабанов Опубликовано 07 марта 2012 года Учительница задаёт школьникам задачу: - У меня 36-й размер обуви, живу я на 7-м этаже и езжу на работу в школу на 42-м троллейбусе.
Дмитрий Шабанов: Вперёд, к эпигенетическому будущему! Дмитрий Шабанов
Дмитрий Шабанов: Вперёд, к эпигенетическому будущему! Дмитрий Шабанов Опубликовано 29 ноября 2012 года В прошлой колонке я похвастался, что в конференции на харьковском биофаке, к организации которой я имел отношение, принимал участие Александр
Дмитрий Шабанов: Невидимая Нога и национальный вопрос Дмитрий Шабанов
Дмитрий Шабанов: Невидимая Нога и национальный вопрос Дмитрий Шабанов Опубликовано 24 октября 2012 года Совпали как-то в последнее время острые дискуссии, связанные с разницей национальностей и культур, идущие и в интеллектуально и культурно близкой