Дмитрий Шабанов: Когда отбор становится неэффективным? Опубликовано 22 февраля 2012 года
Дмитрий Шабанов: Когда отбор становится неэффективным?
Опубликовано 22 февраля 2012 года
sub { font-size: 8pt; }
Да-а-а, судя по отзывам на предыдущую колонку, многих читателей она не убедила. Я имею в виду не креационистов и иже с ними, как заклинание повторяющих, что «факты и логика опровергают эволюцию», и демонстрирующих незнакомство с фактами и неумение пользоваться логикой. Надо бы научиться как-то отвечать таким читателям, чтобы они потеряли интерес к повторению своих бесхитростных аргументов, но дело не в них.
Одна из проблем понимания проявилась в том, что часть читателей решили, что если я говорю о том, что слоны должны эволюционировать медленнее фораминифер согласно СТЭ (синтетической теории эволюции), то скоростная эволюция слонов оказывается чем-то необъяснимым или чудесным. Пожалуйста, не отождествляйте СТЭ и эволюционную биологию вообще! СТЭ с её семидесятипятилетней историей – лишь одна из заслуженных теорий, хоть и весьма авторитетная. Поверьте: современная биология – одна из самых динамичных наук.
Более серьёзная проблема связана с очень конкретным мышлением многих читателей. Я привел умозрительный пример с тремя близнецами, которые росли в разных условиях, и подтолкнул комментаторов к обсуждению жизненных перипетий этих виртуальных людей. А как повлияет на эволюцию слонов то, что они могут ходить с места на место? А как эволюция видов может учитывать то, что ждет их впереди?..
Сосредоточенность на тех или иных житейских историях (бывает так, а бывает и этак...) мешает анализировать влияние отдельных факторов на эволюцию. Как сделать эту связь нагляднее? Связь исходных посылок и вытекающих из них выводов в любой теории не случайна. Не все могут умозрительно представить, как связано изменение жизнеспособностей каких-то категорий особей с их эволюцией. Как сделать для таких читателей мои рассуждения более наглядными?
Построить модель! Модели, даже простые, позволяют понять, какие следствия вытекают из начального набора свойств системы, которые были учтены при её моделировании.
Этим я и займусь. Сила СТЭ (одна из причин, благодаря которой эта теория ещё остается на плаву) в том, что она очень формализована и опирается на математический аппарат генетики популяций. Процессы, протекающие в соответствии с её предсказаниями, легко моделировать! Как обосновать, что моя модель отражает предсказания, которые должны делаться в соответствии с логикой СТЭ? Тут не ограничишься коротким определением СТЭ, от одного из её противников, как в прошлой колонке. А что считать каноном СТЭ? Поскольку у этой теории не было единого автора, её создатели как-то не собрали воедино основные положения своего детища. Формальнее всего описал новую теорию Николай Владимирович Тимофеев-Ресовский (1900-1981), работавший в то время в Германии. В 1939-1940 году он выделил элементарные структуры, материал и факторы эволюции. Первым «скелет» (набор постулатов) СТЭ предложил с полемической целью её яростный противник, Александр Александрович Любищев (1890-1972). Из сторонников СТЭ, кажется, это впервые сделал Николай Николаевич Воронцов (1934-2000), один из учеников Тимофеева-Ресовского.
Воронцов собрал список постулатов СТЭ в 1978-1980 годах. Но и развитие биологии, и собственные исследования (к примеру, роли хромосомных перестроек в видообразовании) со временем убедили Воронцова в ограниченности такого канона (и соответственно ограниченности СТЭ). В конце жизни он описал, в чём его понимание биологии конца прошлого века расходилось со СТЭ. Я имел нескромность выложить на своём сайте таблицу, где сопоставлены постулаты СТЭ по Воронцову и его уточнения к ним, сделанные через 20 лет. Не забывайте, что и правый столбец таблицы по Воронцову – ещё не конец развития эволюционной биологии...
Итак, формализованный характер СТЭ позволяет утверждать, что сделанная мною модель отражает положения этой теории. Модель сделана в Excel, если хотите, можете скачать её на моём сайте; впрочем, надеюсь, что всё будет понятно и из самой колонки. Вообще, Excel дает замечательные возможности для моделирования, делая его доступным для людей без математического и программистского образования.
В модели рассматривается популяция, состоящая из свободно скрещивающихся организмов. Как это и принято в СТЭ, фенотипы (и приспособленность организмов) определяются их генами. Рассмотрим два гена, A и B, каждый из который представлен двумя аллелями: A и a для гена A, B и b для гена B. Итак, в популяции могут существовать девять генотипов: AABB, AABb, AAbb, AaBB, AaBb, Aabb, aaBB, aaBb, aabb.
Слишком просто? И этого минимума хватит, чтобы комбинаторика генов оказалась не вполне тривиальной.
Итак, зададим численность популяции (обозначим её K – ёмкость среды – максимальное количество особей, которое может обитать в данных условиях). Зададим начальные доли генотипов (PAAbb; PAaBB и так далее). Что дальше?
По СТЭ, источник новых признаков – новые гены, возникающие в результате мутаций; не будем пока с этим спорить. Зададим в модели частоты, с которыми происходят переходы одних аллелей в другие: Pa?A, PA?a, Pb?B и PB?b.
Осталось предусмотреть ту силу, которая должна менять соотношение аллелей в определённом направлении, – отбор. Для каждого генотипа укажем его приспособленность (fitness): FAAbb; FAaBB и прочие.
Модель работает так. Исходя из распределения генотипов в популяции, вычисляется состав гамет, которые они будут производить. Для упрощения мы рассматриваем гермафродитные организмы, которые производят и яйцеклетки, и сперматозоиды (один раз в своей жизни). Вероятность встречи гамет с любыми генотипами одинакова (например, так может быть, если гаметы выбрасываются на волю случая в воду, как это делают многие морские донные животные).
При вычислении состава гамет учитывается вероятность мутаций. Комбинации гамет определяют генотипы потомства. Шансы на выживание потомков зависят от их приспособленности.
Всё, цикл работы модели окончен. Осталось определить состав следующего поколения. Округление долей генотипов в популяции до единиц особей носит вероятностный характер. К примеру, величина 1,4 округлится до 2 с вероятностью 0,4, и до 1 — с вероятностью 0,6.
Осталось повторить описанный цикл много раз (модель построена для 500 поколений). Выведем на график самый интересный, с точки зрения СТЭ, показатель – динамику соотношения альтернативных аллелей. Компоненты модели я расположил на листе Excel так, чтобы в один экран попадали и график, и окошки для ввода входных значений (обозначения перечислены выше). Скрины этого экрана и иллюстрируют дальнейшее изложение.
Ну что, начнём.
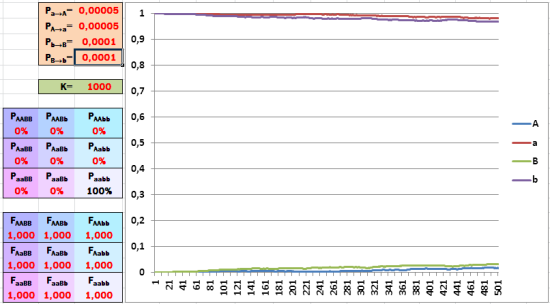
Начальный состав популяции полностью состоит из особей aabb. Приспособленность всех генотипов одинакова, состав популяции постепенно меняется вследствие мутаций (по гену A чуть быстрее, чем по гену B, из-за разной частоты мутирования).
Если в начальном состоянии популяции по каждому гену представлен лишь один аллель, мутации постепенно повышают долю альтернативных аллеей. А что будет, если соотношение аллелей окажется равновесным?
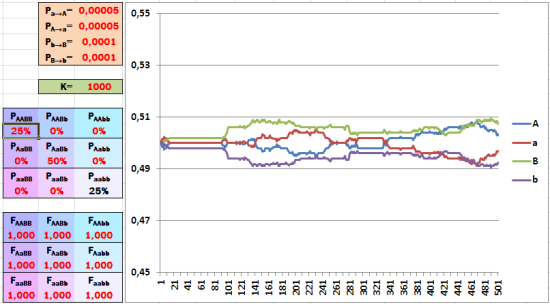
Начальное соотношение аллелей равное и колеблется под влиянием случайностей (мутаций и смещения при дискретизации). Обратите внимание на изменение масштаба по оси ординат.
В отсутствие отбора частоты аллелей начнут «гулять» вокруг среднего значения. Кстати, иногда такие случайные смещения могут привести к потере одного из аллелей.
Пора «включать» отбор. Пусть особи, которые обладают «удачным» генотипом, имеют на 1 процент большие шансов на выживание, чем все прочие. Вам кажется, что такое малое преимущество не может сыграть свою роль в эволюции?
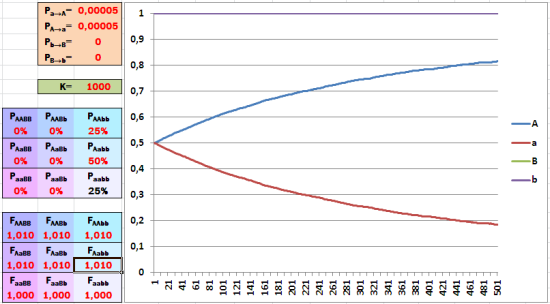
Те же условия, что и в прошлом случае, только включился отбор. Однопроцентного преимущества носителей доминантного аллеля A достаточно, чтобы соотношение аллелей по первому гену быстро менялось (второй пока «выключен» из игры).
Если приспособленность особи определяется единственным геном, преимущества в 1% для носителей доминантного гена достаточно для довольно быстрого изменения соотношения аллелей. А как отразится на скорости эволюции рецессивность аллеля, которому способствует отбор?
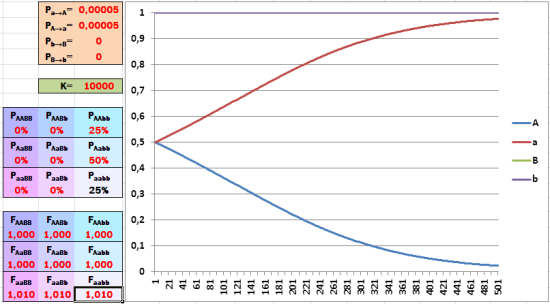
Отбор в пользу рецессивного аллеля (из состояния равенства частот двух альтернатив) идёт ещё эффективнее!
Ускорит (при условии начального равенства частот аллелей). Смотрите: за время, рассмотренное в модели, аллель, которому благоприятствует отбор, почти вытеснил свою альтернативу!
В действительности и популяции, и виды отличаются не по одному, а по множеству генов. В нашей модели мы можем рассмотреть только одновременный отбор по двум из них. Как скажется на скорости эволюции вовлечение второго гена с ещё одной парой аллелей? По гену B тоже поддержим отбором рецессивный аллель (b).
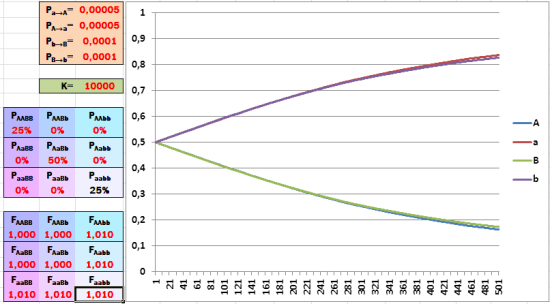
Включаем в «игру» второй ген... и эффективность отбора существенно снижается.
Смотрите, как затормозился отбор! Приспособленность фенотипа особи зависит одновременно от аллелей двух независимых генов; комбинаторика каждого из них мешает отбору по другому гену.
На нашей, достаточно простой модели это не показать, но если на приспособленность особи влияет одновременно 12 генов (по каждому из которых происходит выбор между двумя аллелями), отбор становится совершенно неэффективным! Этот феномен называется дилеммой Холдейна, и обнаружен не кем-нибудь, а одним из создателей СТЭ Дж.Б.С. Холдейном (1892-1964).
Кстати, русская Википедия пишет, что дилемму Холдейна любят креационисты. Напоминаю: это аргумент против СТЭ, а не против эволюции как таковой!
Как мы убедились, при равенстве частот аллелей эффективнее идёт отбор в пользу рецессивного аллеля. Отбор в пользу редкого аллеля эффективнее, если тот доминантен.
Осталось проиллюстрировать одно обстоятельство, затронутое в прошлой колонке. Выставляем в модели высокую поддержку отбором исходно редкого доминантного аллеля – 10%. За 500 поколений отбора такой силы хватает, чтобы редкий аллель стал преобладающим.
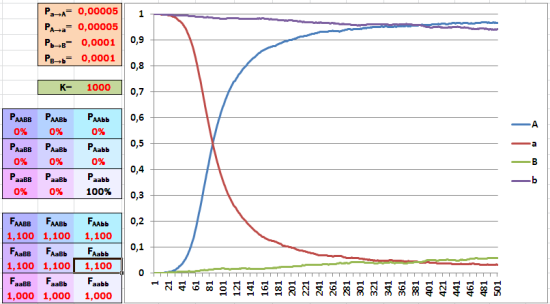
Увеличение доли редкого аллеля идет эффективнее в случае его доминантности.
Но, как мы говорили в прошлый раз, у сложных организмов гены взаимодействуют. Рассмотрим два доминантных аллеля разных генов. Когда они встречаются вместе, они дают значительный выигрыш – в целых 20%. Порознь они дают 1% проигрыша. Предположим, в популяции таких генов очень мало. Сможет ли отбор так же эффективно поднять их долю, как в прошлом примере?
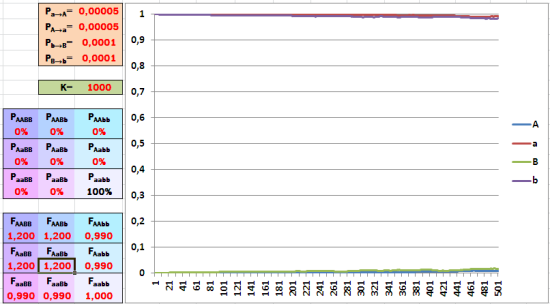
Пример взаимодействия генов. Порознь редкие доминантные аллели двух разных генов дают снижение приспособленности в 1 процент. Зато особи, в которых эти два аллеля встретятся, получат преимущество в 20 процентов. При этих условиях отбор повышает долю таких аллелей крайне медленно.
Увы. Небольшого проигрыша особей, имеющих лишь один из взаимодополняющих генов, достаточно, чтобы этих генов в популяции оставалось немного. Отбор поддержит счастливчиков с удачным сочетанием генов, но большинство их потомков опять получит эти гены порознь. Популяционная эволюция идёт при таких условиях крайне медленно.
Почему? Когда фенотип однозначно отражает генотип, выживание более приспособленных особей эффективно сортирует гены. Если зависимость приспособленности особи от её генотипа усложняется, механизм эволюции по СТЭ начинает работать неэффективно.
На сегодня хватит. Надеюсь, вы убедились: в случаях, когда на приспособленность особи влияет изменчивость многих генов, а тем более если эти гены демонстрируют сложное взаимодействие (а не простое суммирование своих эффектов), скорость изменения частот аллелей серьёзно замедляется.
Как согласовать с этим то, что сложные организмы, демонстрирующие разнообразные взаимодействия их генов, эволюционируют быстрее своих более простых отдалённых родственников? С точки зрения СТЭ это необъяснимо.
Впрочем, на СТЭ развитие науки не остановилось...
К оглавлению
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Специфичный механизм регуляции или отбор? Обсуждение одной гипотезы о механизмах воспроизводства межвидовых гибридов зелёных лягушек Дмитрий Шабанов
Специфичный механизм регуляции или отбор? Обсуждение одной гипотезы о механизмах воспроизводства межвидовых гибридов зелёных лягушек Дмитрий Шабанов Опубликовано 31 мая 2013 Несколько дней назад поспорил я со своим коллегой. Обсуждая одни и те же
Кивино гнездо: Бодрствующее сознание внутри сна Опубликовано 13 января 2012 года
Кивино гнездо: Бодрствующее сознание внутри сна Опубликовано 13 января 2012 года - Вторая часть статьи. Начало читайте здесь.Недавние исследования, проведённые Урсулой Фосс и её коллегами в Боннском университете (Германия), показали, что активность мозга в
Дмитрий Шабанов: Почему есть мужчины и женщины? Дмитрий Шабанов
Дмитрий Шабанов: Почему есть мужчины и женщины? Дмитрий Шабанов Опубликовано 21 апреля 2011 года Я пообещал обсуждать биологические корни нашего поведения, а потом стал писать о другом — об учебниках, о "зелёных"... Исправляюсь.Мы так привыкли, что
Кафедра Ваннаха: Бизнес на сингулярности Опубликовано 26 декабря 2012 года
Кафедра Ваннаха: Бизнес на сингулярности Опубликовано 26 декабря 2012 года О приходе Раймонда «Рэя» Курцвайля (произносится фамилия потомка беженцев из Австрии именно так) в Google Inc. мир узнал из новостных сетей «общего интереса». Трудно сказать, опередили ли они
Голубятня: Гаджеты года. Часть первая Опубликовано 25 декабря 2012 года
Голубятня: Гаджеты года. Часть первая Опубликовано 25 декабря 2012 года Две последние Голубятни уходящего 2012 года я отдам под хит-парад самых потрясающих открытий, какие мне довелось сделать в hardware. Как и полагается, countdown начнем с конца, поэтому сегодня я
Дмитрий Шабанов: Добиологический отбор Дмитрий Шабанов
Дмитрий Шабанов: Добиологический отбор Дмитрий Шабанов Опубликовано 27 сентября 2011 года Да... Обсуждение аргументов, выдвигаемых против эволюционной биологии, свелось к выяснению отношений с комментаторами на сайте «Компьютерры»...Меня попросили
Дмитрий Шабанов: Для чего женщинам быть красивыми? Опубликовано 15 июня 2011 года
Дмитрий Шабанов: Для чего женщинам быть красивыми? Опубликовано 15 июня 2011 года Тело современного человека служит своеобразной картой, правильное прочтение которой <…> позволяет если не однозначно разгадать загадку эволюции человеческой сексуальности,
Василий Щепетнёв: Пророки и угодники — бегство в пустыню Опубликовано 09 февраля 2011 года
Василий Щепетнёв: Пророки и угодники — бегство в пустыню Опубликовано 09 февраля 2011 года "Ревизор", законченный в январе, три месяца спустя уже шёл на сцене Александринского театра. На премьере изволил присутствовать император Николай Павлович. Много смеялся,
Дмитрий Шабанов: Судьба точек роста в системе декоративного образования Дмитрий Шабанов
Дмитрий Шабанов: Судьба точек роста в системе декоративного образования Дмитрий Шабанов Опубликовано 05 июля 2012 года Ф-ф-ф-у-у-у-х! Учебная практика на биостанции, где я провёл последний месяц, закончилась. Прощальный момент был таков: начальник
Кафедра Ваннаха: Торжество образования? Опубликовано 04 октября 2012 года
Кафедра Ваннаха: Торжество образования? Опубликовано 04 октября 2012 года Как следует из доклада Международной организации экономического сотрудничества и развития, Россия нынче не только самая обширная, но и самая просвещённая страна. В ней 51 процент людей
Обзор монитора Acer G246HL Опубликовано 17 декабря 2012 года
Обзор монитора Acer G246HL Опубликовано 17 декабря 2012 года Корпорация Acer выпустила на рынок серию мониторов G6, предназначенных для игровых и офисных приложений, а также для воспроизведения видеоконтента (включая видео высокого разрешения). Основные преимущества
Голубятня: Пятый Опубликовано 18 декабря 2012 года
Голубятня: Пятый Опубликовано 18 декабря 2012 года В минувшую пятницу начались продажи пушистого Пятого Айфона (пушистого, потому что «белого», то бишь официально благославленного Надкусаном) на просторах Отечества. Цены оказались неожиданно низкими в
Дмитрий Шабанов: Реальность vs. действительность Опубликовано 13 декабря 2012 года
Дмитрий Шабанов: Реальность vs. действительность Опубликовано 13 декабря 2012 года Я глубоко убеждён, что есть я на свете, нет меня на свете, а туманность Андромеды летит себе в сторону нашей Галактики. И этой туманности, как, впрочем, и всему остальному миру,
Дмитрий Шабанов: Добиологический отбор
Дмитрий Шабанов: Добиологический отбор Автор: Дмитрий ШабановОпубликовано 27 сентября 2011 годаДа... Обсуждение аргументов, выдвигаемых против эволюционной биологии, свелось к выяснению отношений с комментаторами на сайте "Компьютерры"...Меня попросили развить тему
Дмитрий Шабанов: Когда отбор становится неэффективным?
Дмитрий Шабанов: Когда отбор становится неэффективным? Автор: Опубликовано 22 февраля 2012 годаsub { font-size: 8pt; } Да-а-а, судя по отзывам на предыдущую колонку, многих читателей она не убедила. Я имею в виду не креационистов и иже с ними, как заклинание повторяющих, что "факты и