О природе мужского и женского, или от конъюгации к оогамии Дмитрий Шабанов
О природе мужского и женского, или от конъюгации к оогамии
Дмитрий Шабанов
Опубликовано 17 февраля 2014
Я продолжаю серию «половых» колонок, точнее — колонок об эволюции рекомбинации. Эта — уже восьмая по счету, а вот — первая). После того как я объяснил, почему предполагаю, что половое размножение могло возникать у гермафродитов, я хочу обсудить теорию, которая создаёт серьёзные проблемы для таких представлений. Меня спрашивали, почему я «молчал» (не писал новые колонки) две недели. Не мог уложить в голове, как то, что я изложу здесь, сочетается с тем, о чем писал раньше. Вопрос для меня остаётся открытым. Решение, которое я нашёл, таково: здесь я изложу существенные для меня факты, которые сделают для читателей более понятным разнообразие форм полового процесса, а с главными выводами, которые я пытаюсь сформулировать, повременю.
Я начну с истин из разряда тех, которые излагают в учебниках. Половой процесс связан с оплодотворением — слиянием наследственного материала двух клеток. Как я писал во второй колонке из «половой серии», в типичном случае оплодотворение состоит из двух этапов — слияния двух клеток (сингамии) и слияния их ядер (кариогамии). Что это за клетки, которые сливаются? Это могут быть неспециализированные клетки, представляющие часть тела или все тело одноклеточного организма, — и такой случай можно назвать соматогамией. Чаще всего понятие «соматогамия» применяют только к грибам, у которых могут соединяться клетки из разных гиф, но в более широком смысле его можно применить и ко многим другим случаям, когда для оплодотворения используются клетки, которые выполняют и иные функции. В этой, широкой трактовке самым распространённым типом соматогамии является конъюгация. При ней клетки двух организмов (одноклеточных или многоклеточных) соединяются мостиками, через которые передаются мигрирующие ядра (иногда — вместе со всем клеточным содержимым). При конъюгации информация может передаваться в обе стороны (например, у инфузории туфельки) или только в одну (например, у водоросли спирогиры). Естественно, первый случай аналогичен перекрёстному гермафродитизму, а второй — раздельнополости.
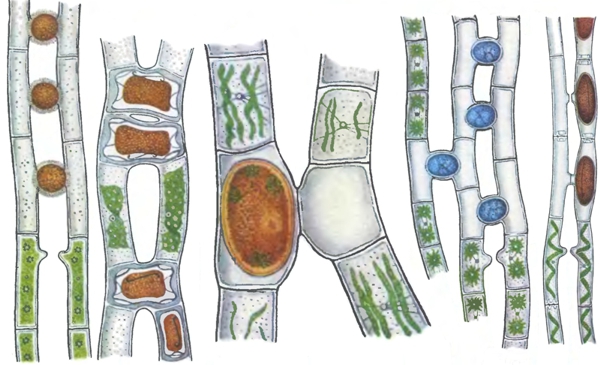
Так происходит конъюгация у некоторых водорослей (рисунок из классического справочника «Жизнь растений»).
Конъюгация (и другие формы соматогамии в широком смысле слова) кажется вам примитивным, архаичным решением? Давайте я расскажу, как она происходит у самых сложных по своему строению клеток, созданных земной эволюцией. Я говорю о наиболее высокоорганизованных одноклеточных — об инфузориях, таких как «школьная» инфузория туфелька.
Размножаются (делятся) инфузории, перешнуровываясь поперёк своей длинной оси. А при конъюгации две инфузории прижимаются друг к другу длинными сторонами.
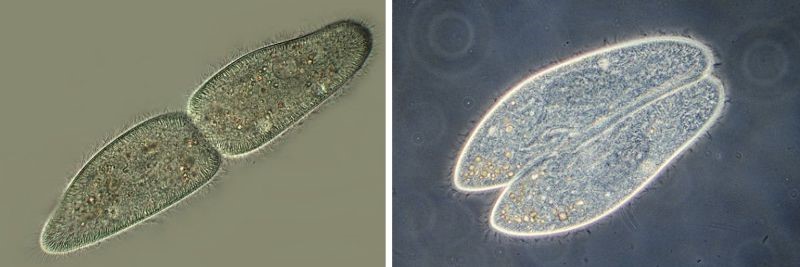
Размножение (слева) и конъюгация (справа) у инфузории туфельки.
Размножение происходит без полового процесса, а половой процесс — без размножения. Другое дело, что после полового процесса инфузории становятся, по сути, новыми индивидами, которые в дальнейшем переходят к делению — клональному размножению. Как ни удивительно, состоящий из многих поколений клеток клон проходит через стадии незрелости, взросления, зрелости и старости. После окончания периода незрелости (в течение которого сменяется некое количество поколений клеток, размножающихся делением; у туфельки он длится около месяца) клон оказывается способен к конъюгации. Она возможна с генетически иными клонами, которые относятся к другому типу спаривания (некоему аналогу пола). Количество типов спаривания у разных инфузорий может быть разным: 2, 4, 6, 7, 8, 10...
Толчком для перехода к половому процессу могут стать ухудшение (с точки зрения инфузории) условий обитания. Пока все идёт хорошо, инфузории делятся, сохраняя соответствующий (раз для его обладателя все идёт хорошо) условиям жизни генотип. Однако если инфузория делилась много сотен раз и при этом не проходила через половое «обновление», то со временем она утрачивает способность к дальнейшим делениям и гибнет.
Представьте себе: в культуре растёт и развивается стареющий клон инфузорий. Раз все индивиды принадлежат к одному клону, они обладают и одним типом спаривания. Начиная с какого-то времени вся экспериментальная популяция оказывается обречена: составляющие её особи не могут найти партнёров для полового процесса. Клон состарился: инфузории живут, делятся, но при любом развитии событий через некое количество поколений их размножение остановится... Парадокс? Безусловно. Здесь проявляется какой-то механизм, не дающий инфузориям отказаться от периодического полового процесса.
Клетка инфузории очень сложна, и необходимые для её жизнедеятельности гены многократно копируются в одном из двух ядер инфузории — большом ядре, макронуклеусе. Перед конъюгацией макронуклеус разрушается, а микронуклеус, малое ядро, проходит через мейоз, делясь на четыре уникальных ядра с уменьшенным вдвое количеством генетической информации. Три из четырёх ядер гибнут, а одно делится ещё раз с помощью митоза, образуя стационарное («женское») и мигрирующее («мужское») ядра. Две инфузории обмениваются мигрирующими ядрами, которые сливаются со стационарными ядрами инфузории-партнёра, образуя новые микронуклеусы. В результате получается два генетически идентичных индивида-эксконъюгата...
Почему я потратил столь много места на обсуждение случая инфузорий? Он показывает, что размножение без образования специфических клеток могут использовать чрезвычайно высокоорганизованные существа. Более того, на их примере прослеживается связь между половым процессом и феноменом смерти. Поверьте, хотя эта связь недостаточно осознана, она весьма глубока. Не тратя пока что времени на её обоснование, выскажу важную мысль: причины, которые обеспечивают половой процесс, тесно связаны с причинами, обеспечивающими запрограммированную смерть индивидов...
Перейдём ко второму, намного более распространённому типу полового процесса — с использованием специализированных половых клеток, гамет. Этот тип полового процесса называется гаметогамией. Гаметы могут производить как одноклеточные, так и многоклеточные организмы. Известны три основные формы гаметогамии. При изогамии гаметы, которые сливаются друг с другом, имеют одинаковый размер. Это подвижные клетки, которые интенсивно ищут друг друга. Эти клетки могут быть совершенно одинаковыми внешне, но при этом они все равно несут биохимические маркеры, обеспечивающие слияние (сингамию) различных клеток. О поле здесь говорить не приходится, разные типы гамет можно обозначить, к примеру, знаками «+» и «—». При анизогамии подвижные гаметы оказываются принадлежащими к двум типам — крупному и мелкому. В данном случае понятно, что крупные гаметы соответствуют женскому полу, а мелкие — мужскому. Логичным завершением этой эволюции оказывается оогамия. Женская гамета теряет активную подвижность и называется яйцеклеткой; её поиск — задача мелких и подвижных гамет, которые называются сперматозоидами.
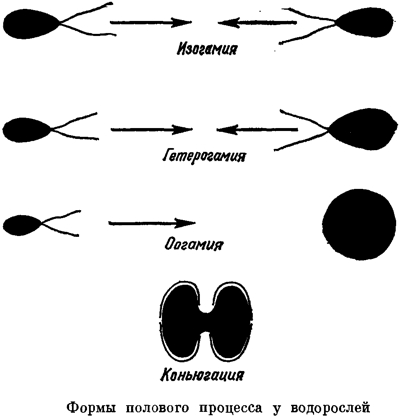
Сравнение трёх форм гаметогамии и конъюгации («Жизнь растений»).
Вероятно, самым простым вариантом гаметогамии является изогамия. Как кажется, этот способ размножения является самым «логичным» и простым. Почему же тогда у большинства высокоорганизованных видов мы наблюдаем оогамию?
Вспомните колонку, в которой я рассказывал о возможном механизме перехода от гермафродитизма к раздельнополости. Успешность того или иного решения зависит не от его оптимальности, а от его устойчивости (или неустойчивости). Изогамия оказывается неустойчивой.
Возможность такого механизма перехода от изогамии к анизогамии, о котором я буду говорить, осознана уже давно. Первая публикация об этом относится к 1972 году (Parker, G. A., Baker, R. R. and Smith, V. G.F. The origin and evolution of gamete dimorphism and the male-female phenomenon. Journal of Theoretical Biology, 1972, v. 36, p. 529–553). Эту концепцию принято называть PBS-теорией, по первым буквам фамилий трёх её авторов. Со времени создания этой теории моделирование взаимодействия гамет разного размера превратилось в отдельную отрасль науки.
Я опишу основную мысль, пользуясь своей моделью, намного более примитивной, чем наилучшие образцы, созданные для изучения этой проблемы. Модель можно скачать с моего сайта: или версию для Excel-2013, с которой я и работал, или немного упрощённый вариант для Excel-2003.
Рассмотрим популяцию организмов, которые выбрасывают гаметы в воду. Начальное состояние — типичная изогамия. Организмы, относящиеся к обоим типам скрещивания (1 и 2), производят одинаковые по размеру гаметы. В том случае, который показан на рисунке, размер всех гамет составляет 12,5% от максимально возможного.
Гаметы встречаются в толще воды, сливаются и образуют зиготу. С вероятностью в 50% особь принадлежит к первому или второму типу. Она будет производить такие же по размеру гаметы, которые производил её родитель, относящийся к её типу скрещивания, с небольшим уточнением. С небольшой вероятностью d (в показанном примере d=0,005) особь переходит к производству гамет соседнего размерного класса: в 2 раза крупнее или в 2 раза мельче. Все особи вкладывают в производство гамет одно и то же количество энергии, и поэтому те особи, которые производят вдвое более мелкие гаметы, выбросят в воду в 2 раза большее их количество.
Последнее, что нужно пояснить перед обсуждением результатов, — то, что вероятность выживания зигот зависит от количества энергии, полученной ими от гамет (попросту от размера). Самый простой вариант зависимости — прямая пропорциональность размера зиготы и её шансов на выживание. Посмотрим, что получится.
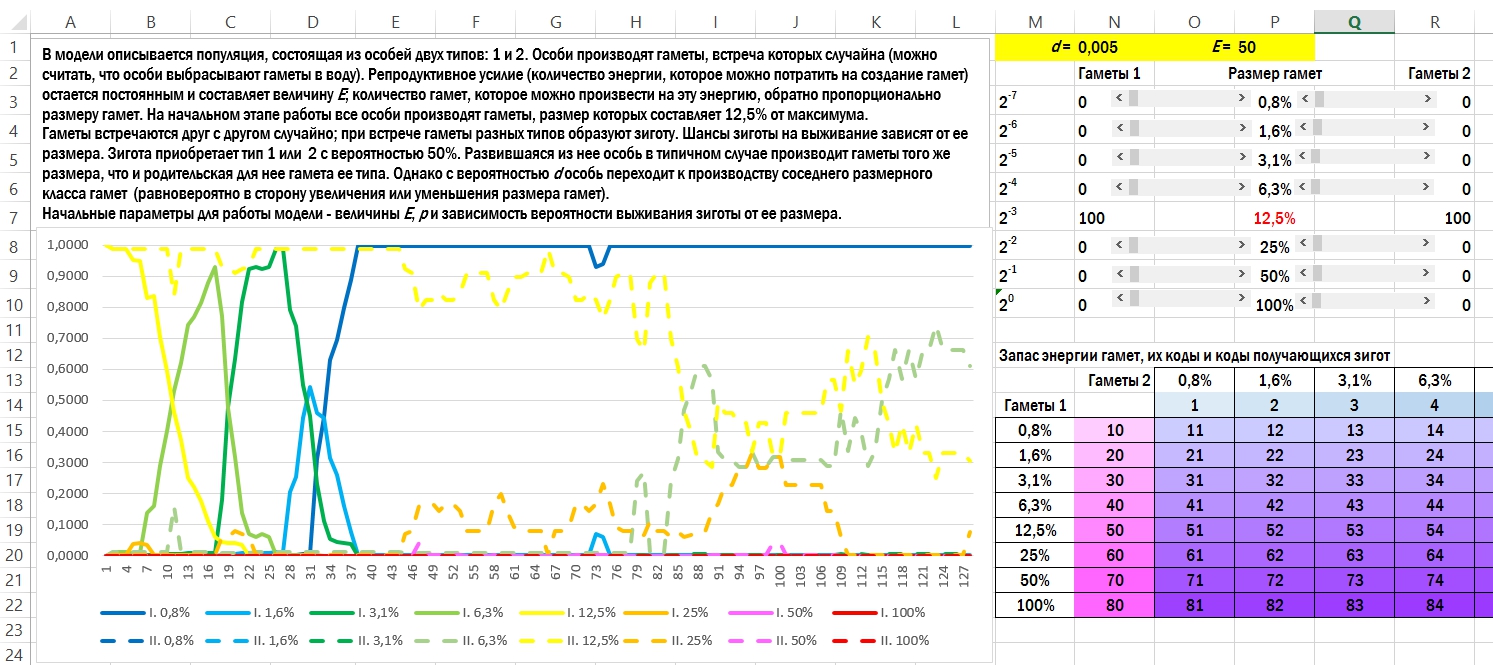
Окно с коротким описанием модели, ячейками для ввода начальных параметров и итоговым графиком её работы. Гаметы первого пола показаны сплошной линией, второго — прерывистой. Выживаемость зигот прямо пропорциональна запасу их энергии. В начальном состоянии оба типа организмов производят гаметы в 12,5% от максимального; к концу имитации первый тип производит исключительно гаметы размером в 1,6%, а второй — в основном в 6,3% от максимального.
Оба «пола» уменьшают размеры своих гамет. Почему — легко понять. При постоянстве размера половых клеток партнёра та особь, которая уменьшит свои гаметы вдвое, вдвое же увеличит количество своих потомков, а их выживаемость сократится только на 75% (запас энергии зиготы зависит от размера обеих гамет). В этой ситуации оба «пола» сокращают размер своих половых клеток. Это происходит до предела, при котором выживаемость зигот становится угрожающе низкой.
Реалистичны ли использованные мной условия? Не вполне. Вероятно, зигота, которая мельче некоего минимального размера, вообще не имеет шансов на выживание. Жизнеспособность зиготы в таком случае должна быть пропорциональна разнице между её размером и этим минимумом.
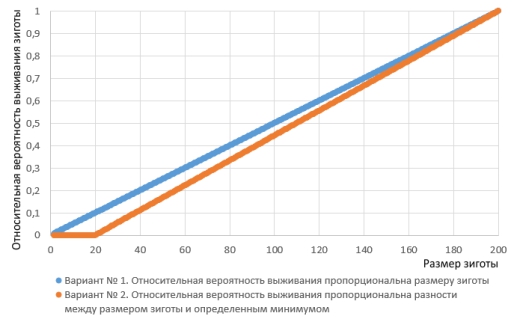
Два варианта зависимости шансов на выживание зиготы от её размера (обусловленного размером гамет).
Стоит перейти с первого варианта зависимости шансов на выживание зиготы ко второму, поведение модели кардинально меняется. Увеличение размеров гамет тоже оказывается выгодной стратегией (приглядитесь: события на картинке внизу начинаются с того, что «пол», показанный пунктиром, переходит к производству более крупных половых клеток).
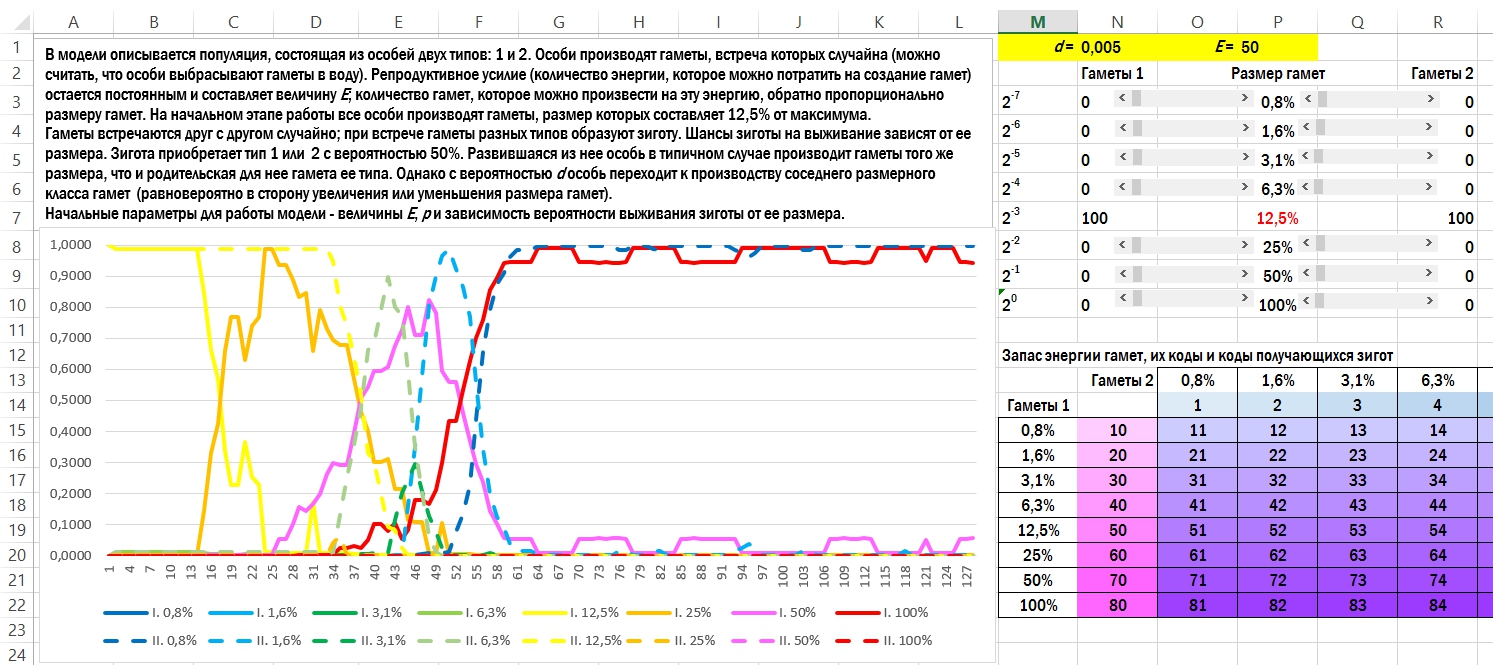
Здесь реализован второй вариант зависимости шансов на выживание зиготы от её размера. В начале имитации оба пола производят гаметы в 12,5% от максимального; к концу (через 128 поколений) первый пол даёт почти исключительно гаметы в 0,8%, а второй — в 100% от максимального размера.
При этих условиях модель быстро переходит к состоянию, где один пол производит самые крупные из возможных гамет, а второй — самые мелкие. Какой именно пол окажется «крупногаметным», а какой — «мелкогаметным», решает случай.
Как видите, в нашей модели мы наблюдали переход от изогамии к анизогамии.
Как я уже сказал, для изучения этого перехода разработаны весьма изощрённые модели, которые учитывают и иные факторы. Например, очевидно, что вероятность встречи гамет зависит от эффективности их плавания, которая, в свою очередь, зависит от их размеров (в частности, влияющих на число Рейнольдса — величину, определяющую характер движения обтекающей гамету жидкости). Учёт таких факторов приводит к более сложным зависимостям, которые, тем не менее, подтверждают общую закономерность: изогамия относительно неустойчива и с большой вероятностью вытесняется анизогамией.
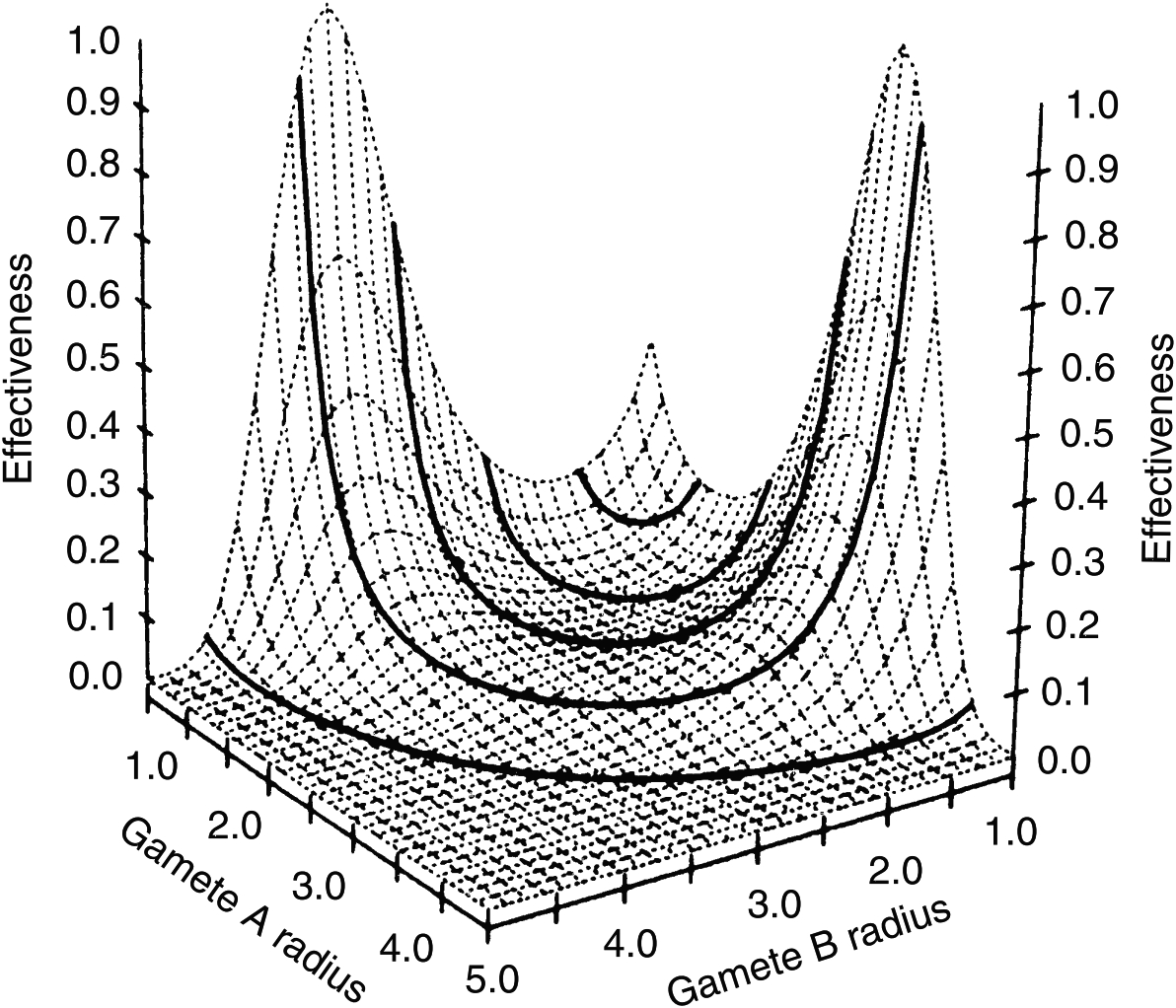
Трёхмерная визуализация результатов намного более сложного моделирования. Высота пиков отражает устойчивость различных сочетаний размеров гамет. Изогамии соответствует небольшой пик посредине, а анизогамии — два симметрично расположенных более высоких пика по краям (из книги: T. Togashi and P.A. Cox (editors). The Evolution of Anisogamy. Cambridge University Press, 2011, 250 p.).
И знаете, что мы в конце концов получили? Раздельнополость! Дело в том, что самцы — это пол, который производит много мелких половых клеток, а самки — пол, который производит немного крупных. Все прочие несовпадения между самками и самцами являются следствиями этого первичного различия!
Теперь, как мне кажется, внимательным читателям восьми «половых» колонок предоставлены ключевые факты и предположения, на основе которых можно судить о возможных сценариях возникновения полового размножения. Этим вопросом мы в следующий раз и займёмся.
К оглавлению
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Ещё раз о природе этноса, а также о разнообразии надсистем Дмитрий Шабанов
Ещё раз о природе этноса, а также о разнообразии надсистем Дмитрий Шабанов Опубликовано 17 ноября 2013 «Понял, что нужно уточнить понятия. Эпифеномен для меня — это когда «они» ведут себя одинаково просто потому, что сами одинаковы…»А.
Дмитрий Шабанов: Фотопериодические реакции Дмитрий Шабанов
Дмитрий Шабанов: Фотопериодические реакции Дмитрий Шабанов Опубликовано 02 ноября 2011 года Мне уже приходилось писать, что меня огорчает недооценка нашей биологической природы. Биологический фундамент, на котором вырастает и наша культура, и
Дмитрий Шабанов: Учебники. Прямиком в послезавтра Дмитрий Шабанов
Дмитрий Шабанов: Учебники. Прямиком в послезавтра Дмитрий Шабанов Опубликовано 07 апреля 2011 года Пока издательства выпускают учебники по госзаказу, доходчивость, качество, мотивирующие способности того или иного учебника будут оставаться
Дмитрий Шабанов: Блуждание глазами по всему небу Дмитрий Шабанов
Дмитрий Шабанов: Блуждание глазами по всему небу Дмитрий Шабанов Опубликовано 23 августа 2011 года Ибо весьма часто случается, что даже и нехристианин знает кое-что о земле, небе и остальных элементах видимого мира, о движении и обращении, даже
Дмитрий Шабанов: Почему есть мужчины и женщины? Дмитрий Шабанов
Дмитрий Шабанов: Почему есть мужчины и женщины? Дмитрий Шабанов Опубликовано 21 апреля 2011 года Я пообещал обсуждать биологические корни нашего поведения, а потом стал писать о другом — об учебниках, о "зелёных"... Исправляюсь.Мы так привыкли, что
Дмитрий Шабанов: Об извращённой любви к природе Дмитрий Шабанов
Дмитрий Шабанов: Об извращённой любви к природе Дмитрий Шабанов Опубликовано 18 мая 2011 года Я писал уже, что из-за активистов-природоохранников мне бывает стыдно называть себя экологом. Если вы составили представление об экологии по материалам
Дмитрий Шабанов: Чудеса полового размножения Дмитрий Шабанов
Дмитрий Шабанов: Чудеса полового размножения Дмитрий Шабанов Опубликовано 29 февраля 2012 года Обсуждение последних «эволюционных» колонок привлекло внимание к теме эволюции эволюции. Некоторым читателям сложно поверить, что разные группы
Дмитрий Шабанов: Цельная модель бытия Дмитрий Шабанов
Дмитрий Шабанов: Цельная модель бытия Дмитрий Шабанов Опубликовано 21 июня 2012 года N.N.: У тебя бывает так: ты заходишь куда-то в комнату и вот вообще не помнишь, зачем пришёл? M.M.: Ну? N.N.: У меня ощущение, что у собак так проходит вся жизнь. (обрывок
Дмитрий Шабанов: Эволюция: биологическая и социальная Дмитрий Шабанов
Дмитрий Шабанов: Эволюция: биологическая и социальная Дмитрий Шабанов Опубликовано 08 ноября 2012 года Некоторым читателям может показаться странным фокус моего внимания в этой колонке. Я хочу разобраться в том, чем социальная эволюция отличается от
Дмитрий Шабанов: «Инстинкт сохранения вида»? Дмитрий Шабанов
Дмитрий Шабанов: «Инстинкт сохранения вида»? Дмитрий Шабанов Опубликовано 07 июня 2012 года Некий йоговский сайт ничтоже сумняшеся утверждает: «Человек, как и любое другое существо на Земле, появляется для того, чтобы обеспечить процветание и
Дмитрий Шабанов: Судьба точек роста в системе декоративного образования Дмитрий Шабанов
Дмитрий Шабанов: Судьба точек роста в системе декоративного образования Дмитрий Шабанов Опубликовано 05 июля 2012 года Ф-ф-ф-у-у-у-х! Учебная практика на биостанции, где я провёл последний месяц, закончилась. Прощальный момент был таков: начальник
Дмитрий Шабанов: «Экологические задачи» для студентов и школьников Дмитрий Шабанов
Дмитрий Шабанов: «Экологические задачи» для студентов и школьников Дмитрий Шабанов Опубликовано 07 марта 2012 года Учительница задаёт школьникам задачу: - У меня 36-й размер обуви, живу я на 7-м этаже и езжу на работу в школу на 42-м троллейбусе.
Дмитрий Шабанов: Вперёд, к эпигенетическому будущему! Дмитрий Шабанов
Дмитрий Шабанов: Вперёд, к эпигенетическому будущему! Дмитрий Шабанов Опубликовано 29 ноября 2012 года В прошлой колонке я похвастался, что в конференции на харьковском биофаке, к организации которой я имел отношение, принимал участие Александр
Дмитрий Шабанов: Невидимая Нога и национальный вопрос Дмитрий Шабанов
Дмитрий Шабанов: Невидимая Нога и национальный вопрос Дмитрий Шабанов Опубликовано 24 октября 2012 года Совпали как-то в последнее время острые дискуссии, связанные с разницей национальностей и культур, идущие и в интеллектуально и культурно близкой
Дмитрий Шабанов: «Чувствую какую-то неизбежность...» Дмитрий Шабанов
Дмитрий Шабанов: «Чувствую какую-то неизбежность...» Дмитрий Шабанов Опубликовано 22 августа 2012 года Брожу ли я по уличному шуму, ем кашу или моюсь по субботам, я вдумчиво обдумываю думу: за что меня считают идиотом. - Игорь ГуберманВы разрешите, я
Дмитрий Шабанов: Жабы. Скороспелые или тугорослые Дмитрий Шабанов
Дмитрий Шабанов: Жабы. Скороспелые или тугорослые Дмитрий Шабанов Опубликовано 18 апреля 2012 года Закончился тринадцатый сезон, в течение которого мы с моими коллегами изучали серых жаб на их нересте. Мне показалось, что некоторые наши промежуточные