Происхождение пола, раздельнополости и гемиклонального наследования. Постановка задачи Дмитрий Шабанов
Происхождение пола, раздельнополости и гемиклонального наследования. Постановка задачи
Дмитрий Шабанов
Опубликовано 24 декабря 2013
Только-только начав сотрудничать с сетевой «Компьютеррой», я написал колонку, где подвёл разговор к теме происхождения раздельнополости. Эта колонка была так-точетвёртой по счёту. Там я обосновывал, почему раздельнополость, широко распространённую среди высокоорганизованных живых организмов, объяснить не просто. В седьмой колонке я пояснил, почему широко известные (и весьма интересные) идеи Вигена Геодакяна не позволяют объяснить происхождение (именно происхождение, а не успех) раздельнополости. В тех статьях я пообещал вскорости продолжить разговор на эту тему... и не выполнил своего обещания. Тогда мне казалось, что осталось лишь чуть-чуть доработать почти готовую модель. Увы, за прошедшие два с половиной года я так и не смог справиться с этой задачей. Но к теме вернуться всё-таки надо, и я это сделаю — начиная с этой, сто тридцатой по счёту колонки. Теперь я постараюсь отработать тему более основательно.
Первый шаг, который надо сделать, — задать классификацию типов популяционного воспроизводства. Обратите внимание: речь идёт не о типах размножения (их классификации разработаны давным-давно). Размножение — это процесс, который идёт на уровне организмов, а на уровне популяций происходит воспроизводство. Однотипное популяционное воспроизводство может осуществляться благодаря принципиально разным способам размножения.
Поясню эту мысль на примерах.
— Амёба протей делится пополам в результате базового типа деления у всех ядерных организмов, митоза. Вначале делится ядро (с образованием двух идентичных дочерних ядер, потом — цитоплазма клетки.
— Мелкий морской многощетинковый червь Ctenodrilus monostylos утратил способность к половому процессу и размножается с помощью архитомии. При этой форме вегетативного размножения тело разделяется на переднюю и заднюю половины, каждая из которых доращивает недостающую часть.
— Скальные ящерицы из рода даревския (названного так в честь описавшего их размножение И. С. Даревского) размножаются путём партеногенеза (устарелое название — «девственное размножение»). Партеногенез, по принятой классификации, — одна из форм полового размножения; перед образованием яйцеклетки хромосомы в клетке-предшественнице удваиваются путём эндомитоза (удвоения хромосом без деления клетки), и образующаяся яйцеклетка может развиваться без оплодотворения.
— У наездников (перепончатокрылых насекомых), заражённых паразитической бактерией вольбахией, бактерии останавливают первое деление мейоза (клеточного деления, в ходе которого образуются половые клетки), приводя к паразитарно индуцированному партеногенезу.
— Серебряные караси во многих водоёмах размножаются путём гиногенеза, при котором икринка, отложенная самкой карася, активируется сперматозоидом любой карповой рыбы. Ядро сперматозоида в икринку не попадает, и развитие потомства обеспечивается ядром яйцеклетки.

Иллюстрации к упомянутым примерам клонального воспроизводства: амёба Amoeba proteus; архитомия полихеты Ctenodrilus monostylos; скальная ящерица Darevskia unisexualis; наездник Trichogramma и его яйцо, заражённое бактериями Wolbachia (светлые точки); серебряный карась Carassius gibelio.
Описанные формы размножения отличаются друг от друга прямо-таки фундаментально. А с точки зрения популяционного воспроизводства они весьма схожи, так как порождают клоны — совокупности генетически идентичных особей. Во всех этих случаях дочерние особи генетически не отличаются от материнских (строго говоря, это утверждение не абсолютно верно, но для дальнейшего изложения это не существенно). Состоящие из клонов популяции принципиально отличаются от популяций с половым воспроизводством. Фактически это «пучки» из независимых клональных линий, способных ветвиться и отмирать. Кстати говоря, функционирование клональных популяций, механизмы, благодаря которым поддерживается их принадлежность к одному виду, да и вообще понимание того, чем является вид у клональных организмов, разработаны современной наукой совершенно недостаточно...
Итак, одни и те же типы воспроизводства могут развиваться на разной основе. Чтобы их сравнивать, их нужно выделить и обозначить. Классификация типов популяций по механизмам их воспроизводства, которую я предлагаю, показана на схеме. Эта классификация является результатом детализации той, что я публиковал несколько лет назад. Однако перед тем, как перейти к её обсуждению, сделаю важное уточнение.
Классификация, которую мы будем обсуждать, касается только популяций с постоянным способом размножения особей. Кроме них, известно множество случаев сложных жизненных циклов или чередования способов размножения. У многих видов половое размножение чередуется с бесполым или размножение с оплодотворением чередуется с партеногенезом. Так, дафнии и тли в течение лета образуют клональные линии партеногенетических самок, а перед уходом на зимовку у них появляется раздельнополое поколение. Самцы и самки производят оплодотворённые яйца, которые дожидаются следующей весны.
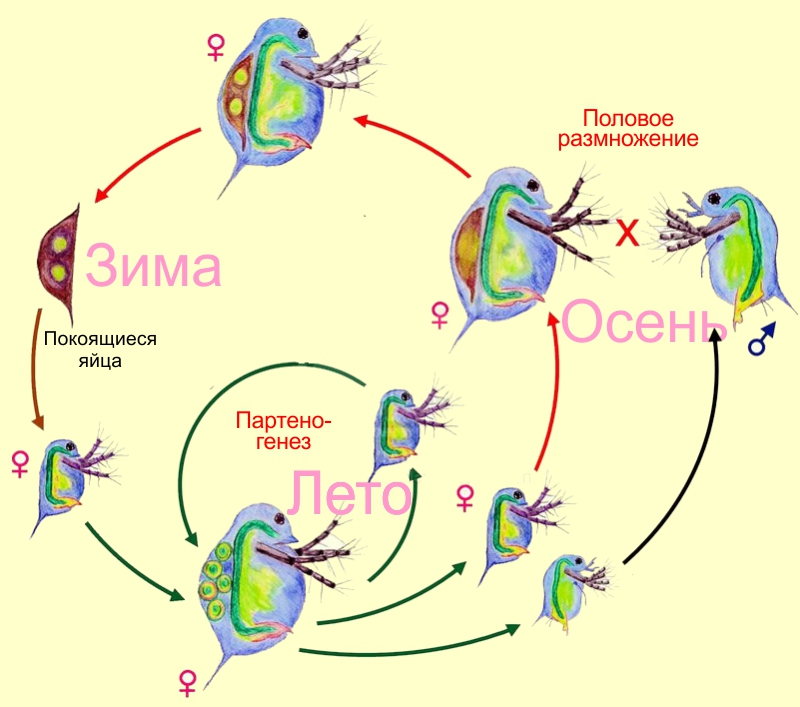
Циклический партеногенез у дафний.
Строго говоря, такие способы воспроизводства не вписываются в предложенную классификацию. Однако всегда можно сказать, к примеру, что дафнии и тли сочетают подтипы воспроизводства IIIa и IIb. Впрочем, чередование поколений — в любом случае отдельный разговор. Отложив его до лучших времён, перейдём к рассмотрению типов популяций с постоянным характером воспроизводства.
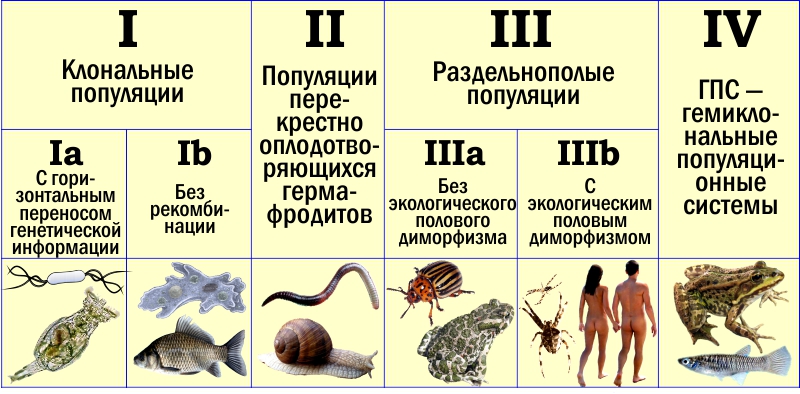
Классификация типов популяционного воспроизводства (у видов с постоянным характером размножения особей). Подробные пояснения — в тексте
Тип I представлен уже упомянутыми клональными популяциями. Он делится на два подтипа в зависимости от того, изолированы клональные линии друг от друга генетически или нет.
Для подтипа Ia характерна способность к горизонтальной передаче генетической информации («вертикальной» считается передача внутри генеалогической последовательности, от предков к потомкам, а «горизонтальной» — между неродственными организмами). Самые разнообразные бактерии относятся именно к этому типу. Горизонтальный перенос генов у прокариот (доядерных организмов) был открыт на примере передачи устойчивости к антибиотикам. В последнее время стало ясно, что такой перенос — широко распространённое явление. Для бактерий нехарактерны изолированные видовые генофонды. «Куски» генетического материала передаются от «вида» к «виду» с помощью различных механизмов.
Именно поэтому в столбце какая-тоIa нарисована бактерия. Под ней находится совершенно иное существо — бделлоидная коловратка. Это микроскопический червь — а следовательно, эукариотический (ядерный) организм. Как у большинства других животных, и в частности у нас с вами, в его клетках находится двойной набор генетической информации, размещённый на двойном (диплоидном) наборе хромосом (содержащих ДНК клеточных структур). Особенность в том, что у нас с вами один набор получен от отца, а другой — от матери, а бделлоидные коловратки утеряли половое размножение в довольно давнем эволюционном прошлом. Наши парные хромосомы содержат сходные (с точностью до индивидуальных отличий) версии одного и того же генетического текста. Парные хромосомы бделлоидных коловраток перестали, по сути, быть парными: они содержат функционально различный текст, который буквально набит фрагментами, полученными от других организмов в результате горизонтального переноса. Размножаются эти удивительные черви путём партеногенеза.
Для бактерий, вероятно, возможна ситуация, когда чужой фрагмент генетической информации встраивается куда попало и оказывается полезен. Такая вставка не является полностью случайным текстом. Это результат отбора, проходившего у других организмов, и он, возможно, связан с какими-то ценными свойствами. Эукариоты перестраивают свою генетическую информацию иначе. Помните, в недавних колонках об эпигенетической теории эволюции я говорил, что развитие высокоразвитых организмов регулируется намного сложнее, чем у бактерий? Если в ходе индивидуального развития происходит многоэтапное взаимодействие между генами и продуктами их работы, если активность генов регулируется сложными генными сетями, то шансов, что случайный кусок генетического текста, встроившийся в случайное место, окажется потенциально полезен, очень немного.
Горизонтальный перенос встречается и у эукариот, но в целом для них характерен совсем иной механизм рекомбинации, тесно связанный с половым размножением. При оплодотворении возникает клетка с удвоенным набором хромосом. Значит, в процессе образования половых клеток набор хромосом должен сократиться вдвое. Это происходит при делении, которое называется мейозом. В мейозе парные хромосомы узнают друг друга и обмениваются фрагментами в ходе процесса, названного кроссинговером (или попросту перекрёстом). Новая генетическая информация, которая несёт следы действия отбора, происходившего у других организмов, попадает при этом не куда попало, а именно в то место хромосомы, где уже имеются механизмы регуляции её активности.
Однако сейчас мы говорим о клональных популяциях, для которых половой процесс и связанная с ним рекомбинация нехарактерны. Мы выделяем подтип IIa, который характеризуется отказом от систематического использования горизонтального переноса. Представители такого подтипа регулярно возникают от предков, относившихся к следующим типам, но, вероятно, могут также происходить и от популяций Ia. На схеме показаны амёба протей и серебряный карась, размножение которых мы уже успели обсудить.
К типу II относятся популяции перекрёстно оплодотворяющихся гермафродитов. На схеме изображены дождевой червь и виноградная улитка: это те животные, что при размножении образуют пары из гермафродитных особей, каждая из которых передаёт своему партнёру мужские половые продукты. Такие существа уже в полной мере используют ключевые преимущества полового размножения и связанной с ним рекомбинации (следствия кроссинговера во время образования половых клеток), но ещё не несут груза раздельнополости.
Тип III. Раздельнополые. Подразделён на два подтипа в зависимости от того, имеются значимые экологические различия между самками и самцами или нет. Может быть, разделять этот тип на подтипы и не стоило... Главная причина, по которой это всё-таки сделано, такова. Есть серьёзные причины, в силу которых экологический половой диморфизм оказывается выгоден, и тут я вполне согласен с Вигеном Геодакяном. Однако очевидно, что раздельнополость должна вначале попросту возникнуть (подтип IIIa) — и лишь потом между двумя полами могут накопиться серьёзные различия, не относящиеся непосредственно к сфере размножения (подтип IIIb).
Подтип IIIa на схеме проиллюстрирован изображениями колорадского жука и зелёной жабы. Кстати, определённый половой диморфизм у зелёных жаб есть: самки у них (как и у многих других гадов) крупнее самцов. Тем не менее образ жизни обоих полов жаб примерно одинаков; они помещены в IIIa именно для того, чтобы показать, что определённые различия между полами могут наблюдаться и в этом подтипе. К подтипу IIIb на схеме отнесены пауки крестовики и люди.
Наконец, тип IV. Гемиклональные (полуклональные) популяционные системы, ГПС. Обычно их не рассматривают, как нечто стоящее особняком от всех остальных типов популяций (само понятие ГПС мы с моими коллегами придумали и ввели всего несколько лет назад). На схеме — съедобная лягушка (Pelophylax esculentus) и американская карпозубая рыба из рода Poeciliopsis, у которого впервые был описан тип размножения, характерный и для лягушек. У таких организмов межвидовые гибриды передают в половые клетки геном одного из родительских видов как единое целое (почти как единое целое). Межвидовые гибриды обитают совместно с представителями родительских видов, скрещиваются с ними и совместно участвуют в воспроизводстве. Из поколения в поколение в таких системах передаются как клональные (практически не изменяющиеся), так и рекомбинантные, обычные геномы.
Исчерпан ли перечень типов воспроизводства? Не знаю. С учётом тех степеней свободы, по которым разнятся перечисленные типы, каких-то иных их сочетаний я не знаю, но это не означает, что их не может быть...
В размещённой ниже таблице показаны результаты сравнения разных типов популяций по тем признакам, которые представляются мне существенными. Обратите внимание на то, что IV тип оказывается вполне самостоятельным. Наименьшие различия вполне закономерно наблюдаются между подтипами IIIa и IIIb. Увы, в этой колонке я успею обсудить не все графы таблицы: к примеру, разговор о рекомбинации потребует отдельной колонки. Всё же надеюсь, что направление для обсуждения я задал.
Тип популяции
Ia
Ib
II
IIIa
IIIb
IV
Генеалогия
Линейная
Сетчатая
Линейная
«Двойная цена» пола
Нет (потомство непосредственно производят все особи)
Есть (потомство непосредственно производят только самки)
Половой отбор
Нет
Малоэффективный
Эффективный
Есть
Уникальность особи
Нет
Наследуемая
Ненаследуемая
Генофонд
Совокупность клонов
Рекомбинантный генофонд популяции
Рекомбинантный генофонд + полуклоны
Изоляция
Поток генов между клонами
Изолированные клоны
Поток генов внутри вида, изолированного от других видов
Поток генов внутри и между видами
Рекомбинация
Хаотичная
Нет
Упорядоченная внутривидовая
Упорядоченная внутри- и межвидовая
А теперь я сделаю то, ради чего, по сути, и писал эту колонку. Я повторю ту схему, что вы уже видели, добавив в неё условные обозначения переходов между типами, которые происходили в ходе эволюции. Непрерывные стрелки обозначают те эволюционные переходы, относительно которых мы знаем, что они происходили, или можем это предполагать с высокой степенью уверенности (например, переход, показанный зелёным цветом, происходил или так: I?II, или так: I?IIIa).
![]()
Я планирую в ближайшее время обсудить логику следующих переходов.
Возникновение полового размножения: I?II (более вероятно) или I?IIIa (зелёные стрелки).
Возникновение раздельнополости: II?IIIa (более вероятно) или I?IIIa (красные стрелки).
Экологическая дифференциация полов: IIIa?IIIb (фиолетовая стрелка).
Отказ от полового процесса и переход к клональному воспроизводству: III?I или II?I (коричневая стрелка).
Возникновение гемиклональности: III?IV (а чем чёрт не шутит? Может, и II?IV) — синяя стрелка.
Возможный отказ от гемиклональности: IV?III (IV?II)— голубая пунктирная стрелка.
Задача этой колонки состояла в том, чтобы задать схему, позволяющую организовать обсуждение только что перечисленных проблем. Я надеюсь, что просто смогу повторить рисунок со стрелочками и с помощью ссылки отправить за пояснениями всех, кому это нужно, сюда, в эту колонку.
Согласны? Поверьте, тут есть о чём рассказать и о чём задуматься.
К оглавлению
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Хватит ли восьми косвенных гипотез, описывающих преимущества полового размножения, чтобы объяснить возникновение пола? Дмитрий Шабанов
Хватит ли восьми косвенных гипотез, описывающих преимущества полового размножения, чтобы объяснить возникновение пола? Дмитрий Шабанов Опубликовано 11 января 2014 Парадоксальность полового размножения, о которой шла речь в моей предыдущей
Дмитрий Шабанов: Фотопериодические реакции Дмитрий Шабанов
Дмитрий Шабанов: Фотопериодические реакции Дмитрий Шабанов Опубликовано 02 ноября 2011 года Мне уже приходилось писать, что меня огорчает недооценка нашей биологической природы. Биологический фундамент, на котором вырастает и наша культура, и
Дмитрий Шабанов: Учебники. Прямиком в послезавтра Дмитрий Шабанов
Дмитрий Шабанов: Учебники. Прямиком в послезавтра Дмитрий Шабанов Опубликовано 07 апреля 2011 года Пока издательства выпускают учебники по госзаказу, доходчивость, качество, мотивирующие способности того или иного учебника будут оставаться
Дмитрий Шабанов: Блуждание глазами по всему небу Дмитрий Шабанов
Дмитрий Шабанов: Блуждание глазами по всему небу Дмитрий Шабанов Опубликовано 23 августа 2011 года Ибо весьма часто случается, что даже и нехристианин знает кое-что о земле, небе и остальных элементах видимого мира, о движении и обращении, даже
Дмитрий Шабанов: Почему есть мужчины и женщины? Дмитрий Шабанов
Дмитрий Шабанов: Почему есть мужчины и женщины? Дмитрий Шабанов Опубликовано 21 апреля 2011 года Я пообещал обсуждать биологические корни нашего поведения, а потом стал писать о другом — об учебниках, о "зелёных"... Исправляюсь.Мы так привыкли, что
Дмитрий Шабанов: Об извращённой любви к природе Дмитрий Шабанов
Дмитрий Шабанов: Об извращённой любви к природе Дмитрий Шабанов Опубликовано 18 мая 2011 года Я писал уже, что из-за активистов-природоохранников мне бывает стыдно называть себя экологом. Если вы составили представление об экологии по материалам
Обсуждение перехода от гермафродитизма к раздельнополости как пример неклассического развития естественнонаучной гипотезы Дмитрий Шабанов
Обсуждение перехода от гермафродитизма к раздельнополости как пример неклассического развития естественнонаучной гипотезы Дмитрий Шабанов Опубликовано 02 февраля 2014 Я рад тому, как была принята позапрошлая колонка (не прошлая, политическая, а
Дмитрий Шабанов: Чудеса полового размножения Дмитрий Шабанов
Дмитрий Шабанов: Чудеса полового размножения Дмитрий Шабанов Опубликовано 29 февраля 2012 года Обсуждение последних «эволюционных» колонок привлекло внимание к теме эволюции эволюции. Некоторым читателям сложно поверить, что разные группы
Дмитрий Шабанов: «Инстинкт сохранения вида»? Дмитрий Шабанов
Дмитрий Шабанов: «Инстинкт сохранения вида»? Дмитрий Шабанов Опубликовано 07 июня 2012 года Некий йоговский сайт ничтоже сумняшеся утверждает: «Человек, как и любое другое существо на Земле, появляется для того, чтобы обеспечить процветание и
Дмитрий Шабанов: Судьба точек роста в системе декоративного образования Дмитрий Шабанов
Дмитрий Шабанов: Судьба точек роста в системе декоративного образования Дмитрий Шабанов Опубликовано 05 июля 2012 года Ф-ф-ф-у-у-у-х! Учебная практика на биостанции, где я провёл последний месяц, закончилась. Прощальный момент был таков: начальник
Дмитрий Шабанов: «Экологические задачи» для студентов и школьников Дмитрий Шабанов
Дмитрий Шабанов: «Экологические задачи» для студентов и школьников Дмитрий Шабанов Опубликовано 07 марта 2012 года Учительница задаёт школьникам задачу: - У меня 36-й размер обуви, живу я на 7-м этаже и езжу на работу в школу на 42-м троллейбусе.
Первые, вторые и третьи репликаторы по Сьюзен Блэкмор, происхождение жизни и общая схема фазовых переходов в эволюции Дмитрий Шабанов
Первые, вторые и третьи репликаторы по Сьюзен Блэкмор, происхождение жизни и общая схема фазовых переходов в эволюции Дмитрий Шабанов Опубликовано 07 марта 2013 Наэлектризовавшую читателей тему двух предыдущих колонок не затрагиваю; поговорю здесь
Дмитрий Шабанов: Невидимая Нога и национальный вопрос Дмитрий Шабанов
Дмитрий Шабанов: Невидимая Нога и национальный вопрос Дмитрий Шабанов Опубликовано 24 октября 2012 года Совпали как-то в последнее время острые дискуссии, связанные с разницей национальностей и культур, идущие и в интеллектуально и культурно близкой
Дмитрий Шабанов: Последний концерт Кондрашина Дмитрий Шабанов
Дмитрий Шабанов: Последний концерт Кондрашина Дмитрий Шабанов Опубликовано 15 октября 2012 годаНа торрент-трекере ***.com произошел сбой RAID-массива. После восстановления его работы в комментариях к странице, посвящённой записи Кирилла Кондрашина, появился следующий текст. По
Эволюция эволюции: от генетического наследования к интратехнической репликации т-мемов Дмитрий Шабанов
Эволюция эволюции: от генетического наследования к интратехнической репликации т-мемов Дмитрий Шабанов Опубликовано 30 марта 2013<p Человек – не статический центр мира, как он долго полагал, а ось и вершина эволюции, что много прекраснее. Пьер Тейяр де ШарденВ этой
Дмитрий Шабанов: "Экологические задачи" для студентов и школьников
Дмитрий Шабанов: "Экологические задачи" для студентов и школьников Автор: Дмитрий ШабановОпубликовано 07 марта 2012 годаУчительница задаёт школьникам задачу: - У меня 36-й размер обуви, живу я на 7-м этаже и езжу на работу в школу на 42-м троллейбусе. Спрашивается: сколько мне