Краткое изложение эпигенетической теории эволюции, или ЭТЭ для занятых Дмитрий Шабанов
Краткое изложение эпигенетической теории эволюции, или ЭТЭ для занятых
Дмитрий Шабанов
Опубликовано 08 декабря 2013
Две недели прошли у меня под знаком сопереживания потрясениям в украинской политике и споров об эпигенетической теории эволюции. О политике — не сейчас; здесь мы обсудим ЭТЭ. Я регулярно слышу упрёки в отсутствии её короткого изложения. Эта колонка — моя попытка такого изложения, учитывающая свежий опыт обсуждения ЭТЭ на сайте КТ, в ЖЖ Александра Маркова, на моем сайте, на заседании клуба «Эволюция» в Киеве (вот расширенная презентация моего доклада), на круглом столе с Сергеем Ястребовым, проведённом во время молодёжной конференции харьковского биофака.
Да, не забудьте:
— здесь не оспаривается факт эволюции, а идёт речь о проблемах изучения её механизмов;
И ещё. Уточним термины.
Наука (частная) — развивающийся комплекс представлений об определённом аспекте действительности, технологий его изучения и изменения, который может включать в себя различные, в том числе частично противоречащие друг другу, гипотезы и теории.
Теория — целостная система взглядов, в которой одни положения выводятся из других. Теорией может стать по мере своего развития гипотеза, объясняющая определённые феномены и обладающая прогностической ценностью.
Эволюция — необратимые изменения биосистем на протяжении исторического времени биосферы. Приводит к изменению существующих биосистем, в том числе их усложнению, повышению их адаптированности к среде, росту их устойчивости, возникновению у них новых свойств и появлению новых типов биосистем. Эволюция — многоуровневый процесс; эволюционируют популяции, виды, надвидовые группы, а также сообщества и экосистемы.
Эволюция эволюции — изменения механизмов эволюции по мере эволюции биосистем.
Эволюционная биология — наука, изучающая механизмы эволюции. Изучение того, как именно шла эволюция, тоже часто включают в сферу компетенции этой науки, но тут она трактуется в узком, указанном смысле.
Онтогенез — индивидуальное развитие особи, совокупность её закономерных и случайных преобразований на протяжении её жизни.
Отбор — преимущественное воспроизводство особей и их групп, зависящее от их свойств; отбор преимущественно сохраняет и воспроизводит более адаптивных особей.
Адаптивность — соответствие организма той среде, в которой он развивается, позволяющее успешно пройти онтогенез и оставить потомков.
Итак, как вы поняли, эволюционная биология — наука, которая включает множество теорий. Её развитие не закончено, и полной картины механизмов многоуровневого процесса эволюции на сегодня нет. Рассматривая историю эволюционной биологии, мы можем увидеть, что в ней то распространялись идеи, принимаемые большинством учёных, то наступали времена разнобоя во мнениях. Чтобы их описать, я воспользуюсь схемой, предложенной Н. Н. Воронцовым (заполняя последнюю строку таблицы, забегу вперёд, отразив то, к чему прихожу в этой колонке).

Одной из теорий, появившихся в эпоху кризиса III синтеза, стала ЭТЭ. Она основывается на результатах, полученных в 1940-х и 1950-х годах советским зоологом И. И. Шмальгаузеном (теория стабилизирующего отбора) и английским генетиком К. Х. Уоддингтоном (эпигенетический ландшафт и «генетическая ассимиляция модификаций»). Основы ЭТЭ сформулировал московский палеонтолог М. А. Шишкин в работах, опубликованных с 1984 по 1988 год. Вклад в разработку теории внесли также его коллеги А. П. Расницын (метафора адаптивного компромисса) и А. С. Раутиан (эволюция как поддержание устойчивости).
Описывая ЭТЭ, её сравнивают со СТЭ, синтетической теорией эволюции, имея в виду именно относительно целостную теорию, сложившуюся к середине XX века. Почему? Современная эволюционная биология — довольно рыхлый и в определённой степени внутренне противоречивый комплекс концепций. Каждая из них более или менее объясняет какой-то комплекс факторов, игнорируя иные данные. Однако СТЭ в силу своей простоты до сих пор остаётся версией «по умолчанию»: это её излагают в школе и вузах, пытаясь отождествить с современной эволюционной биологией целиком.
Настало время дать краткое описание ЭТЭ.
Эпигенетическая теория эволюции рассматривает эволюцию как процесс смены одних стабилизированных путей онтогенеза другими. У представителей высокоорганизованных групп результат онтогенеза обусловлен чрезвычайно сложным комплексом факторов и результатами взаимодействия их эффектов.
На онтогенез влияет взаимодействие следующих факторов и их эффектов:
наследственных задатков:
генетических (последовательностей нуклеотидов в нуклеиновых кислотах — НК);
эпигенетических (химических и пространственных модификаций макромолекул НК);
других (связанных с организацией цитоскелета, набором РНК и регуляторных молекул, конформацией белков и т. д.);
разнообразных средовых влияний;
случайности.
Результат онтогенеза невозможно предсказать однозначно. Его можно охарактеризовать лишь распределением вероятностей различных исходов, среди которых следует выделить норму (состояние, которое поддерживается стабилизирующим отбором) и разнообразные морфозы (уклонения, аберрации). Метафорой, описывающей распределение возможных итогов онтогенеза, является эпигенетический ландшафт К. Х. Уоддингтона. С этой точки зрения возможные онтогенетические пути можно описать как совокупность стабилизированных участков (креодов), точек бифуркации и разделяющих креоды совокупностей маловероятных и неустойчивых состояний.
Стабилизирующий отбор — преимущественное сохранение и воспроизводство особей, онтогенез которых привёл к норме, — повышает устойчивость развития нормы (повышает его вероятность). Эта устойчивость растёт и благодаря росту эквифинальности развития нормы (способности к реализации нормы у всё более различающихся особей), и благодаря повышению автономности такого развития (способности реализовывать норму во всё более разных условиях среды). Это обеспечивается благодаря тому, что отбор перестраивает систему управления онтогенезом в целом (и генотип в частности). В модели эпигенетического ландшафта действие стабилизирующего отбора выглядит как углубление соответствующего креода.
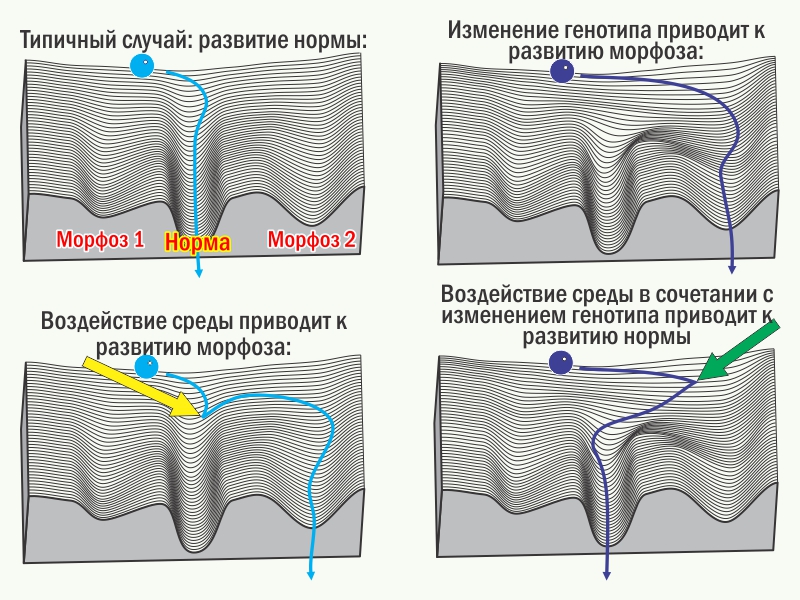
Если характер отбора изменяется и он перестаёт поддерживать прежнюю норму, её развитие дестабилизируется и проявляются разнообразные морфозы. Если какой-то из морфозов оказывается адаптивным, отбор избирательно сохраняет те системы управления онтогенезом, которые привели к такому адаптивному состоянию. Потомки подобных особей с большей вероятностью окажутся адаптивными, если их онтогенез приведёт к тому же результату. Значит, отбор будет поддерживать тех потомков, у которых развитие адаптивного в данных условиях состояния будет оказываться всё более устойчивым (всё более вероятным). Результатом становится повышение устойчивости развития поддерживаемого отбором морфоза, то есть рост его наследуемости. Итак, сам феномен наследственности оказывается результатом отбора.
условия развития. Если отбор поддерживает такой морфоз, он становится новой нормой, его развитие автономизируется и приобретает независимость от специфичных внешних воздействий.
По мере эволюции происходит усложнение системы управления онтогенезом и совершенствование механизмов, обеспечивающих поиск адаптивных морфозов при изменении характера отбора. Возникновение генетического наследования, полового размножения, культурного наследования, сложной социальной организации — некоторые этапы этого процесса.
Чем описанный подход отличается от подхода СТЭ? Для СТЭ эволюция — это перестройка генотипа вследствие отбора по результатам активности генов, отражённым в признаках организма. Именно поэтому СТЭ пытается описать онтогенез как совокупность относительно независимых причинно-следственных связей. Любые взаимодействия, усложняющие передачу информации от генотипа в фенотип, для СТЭ просто помехи, затрудняющие перестройку генотипа по его фенотипическим эффектам.
Если информация от генотипа отражается в фенотипе напрямую, механизм СТЭ работает вполне удовлетворительно. Уравнение Харди — Вайнберга описывает, как соотношение аллелей (альтернативных версий одного гена) у потомков зависит от такового у предков. Математический аппарат СТЭ основан на том, что каждый аллель вносит постоянный вклад в итоговую приспособленность своего обладателя. Если аллель повышает приспособленность организма, отбор будет повышать его частоту и со временем благоприятный аллель вытеснит свои альтернативы. Есть случаи, когда такая модель работает. Рассмотрим два штамма бактерий. Один растёт быстрее, но неустойчив к антибиотику. Другой — устойчив и платит за это более медленным ростом. Их признаки однозначно отражают их генотип. Их динамика на средах с разным содержанием антибиотиков хорошо описывается уравнениями отбора по СТЭ.
Если модель СТЭ верна, эволюция эволюции должна приводить к тому, что фенотип будет все эффективнее, все «прозрачнее» отражать генотип. У таких организмов отбор станет перестраивать генотип особенно эффективно. Они начнут быстрее вырабатывать приспособления и получат преимущество в изменчивой среде. А вот организмы со сложными взаимодействиями разных факторов в онтогенезе должны эволюционировать медленно. Особо «тяжело» — виды, состоящие из долгоживущих и малоплодовитых особей. Оправдывается ли это предсказание?
Нет! Я писал об этом, помните?
В том мире, который мы наблюдаем, самые сложные, самые быстро эволюционирующие с точки зрения изменений своего строения и своего поведения организмы оказываются совершенно невероятными с точки зрения СТЭ. Возьмём, к примеру, людей. Наш генотип содержит совсем немного информации, помещаясь в незаархивированном виде на CD-диск. Собственно гены составляют его небольшую часть; около 25 тысяч наших генов требуют для записи менее 10 мегабайт (а архиватор ужмёт их намного сильнее). Напомню известную шутку, что файл с детальным описанием формы коленной чашечки (одной из самых простых наших костей) в AutoCad займёт больше места. Информация, необходимая для описания нашего строения, несоизмеримо больше ёмкости нашего генома. Ещё удивительнее массив информации в нашей психике. Это означает, что в нашем онтогенезе происходит головокружительное количество выборов возможных путей развития с запоминанием их результата.
Типичным путём эволюционных изменений оказывается для ЭТЭ тот, который соответствует логике опытов Г. К. Уоддингтона. Уоддингтон вызывал у экспериментальных животных морфозы с помощью внешних воздействий. Потомство от скрещивания носителей морфозов опять подвергалось аналогичным воздействиям, и опять для размножения отбирали носителей того же морфоза. Через небольшое (первые десятки) количество поколений эти морфозы начинали развиваться без специфичных воздействий. Неустойчивый, зависимый от среды путь развития становился устойчивым. Чтобы убедиться, что речь не идёт о наследовании приобретённых признаков, сравните эти эксперименты с классическими опытами Вейсмана.
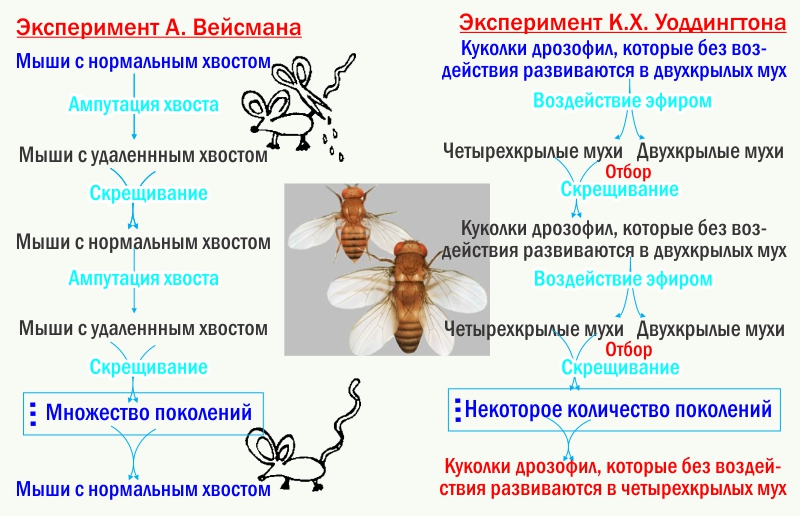
На языке СТЭ опыты Уоддингтона описываются натужно. Отбор на способность развивать определённую модификацию (ненаследуемый признак) привёл к изменению множества генов-модификаторов и в конечном итоге к «генетической ассимиляции модификации», передаче управления развития этим признаком в генотип. Это объяснение предусматривает некие гены-модификаторы, не найденные геномикой. Оно предусматривает стремительную эволюцию этих модификаторов, не соответствующую моделям отбора, разработанным в самой СТЭ. Если принять, что речь идёт не о виртуальных модификаторах, а о других структурных генах, становится непонятно, почему «генетическая ассимиляция» контроля развития одного признака не приводит к хаосу в развитии других.
На языке ЭТЭ опыты Уоддингтона описываются просто. какие-тоПоддерживая морфоз, стабилизирующий отбор приводит к повышению устойчивости его развития. И — обратите внимание! — никаких натянутых попыток представить организм как сумму признаков, а генотип — как сумму генов. Опыты Уоддингтона описывают не превращение «ненаследственных» признаков в «наследственные», а влияние отбора на устойчивость развития. И, кстати, пример людей показывает, что часто «ненаследственные» признаки (то, чему мы учимся) не менее важны для нас, чем иные.
Теперь можно обсудить разнообразие оценок ЭТЭ. Они очень разные. Один полюс заключается в том, что ЭТЭ — полная альтернатива СТЭ, и никакой компромисс между ними невозможен. На другом полюсе находятся те, кто объявляет ЭТЭ лженаукой и даже пытается запретить упоминание имён её сторонников в присутствии студентов, чтоб не заронить сомнения в безальтернативной истинности СТЭ. Поверьте, пишу об таких попытках не умозрительно, а на основе печального опыта... Моя оценка ближе к первому полюсу, хотя отличается от него. Я считаю, что ЭТЭ может стать ядром III синтеза: потенциал для объяснения того, что происходит на организменном уровне, есть только у этой теории.
Конечно, в ЭТЭ сохраняется множество недостаточно разработанных вопросов. Один из них — описание разнообразия признаков с точки зрения регуляции их развития в онтогенезе. Вероятно, даже у сложных организмов есть относительно простые признаки, почти однозначно зависящие от состояния отдельных аллелей. В этих случаях модели СТЭ будут описывать эволюцию таких признаков относительно адекватно. Поломки нормальных путей развития, вероятно, регулируются проще, чем срабатывание уже имеющихся генных механизмов в тех тканях, где они обычно работают. Но возникновение принципиально новых признаков такими механизмами не объяснить...
Итак, по моему мнению, ЭТЭ — более широкое обобщение, чем СТЭ, и случаи, когда подход СТЭ оказывается применим, можно рассматривать (по удачному выражению С. Ястребова), как вырожденные (упрощённые) случаи применимости ЭТЭ.
Важным преимуществом ЭТЭ является, по моему мнению, её способность объяснять быстрое (по эволюционным меркам) появление адаптивных новшеств, гармонично вписывающихся в комплекс признаков организма. Для СТЭ новые адаптивные признаки — результат счастливой случайности, мутации структурного или регуляторного гена, которая оказалась полезной. Чем сложнее организм, тем реже должны происходить такие случайности. Для ЭТЭ новые признаки возникают как ответ целостного организма на изменившиеся условия его развития. В формировании этого ответа отражается весь опыт предшествовавшей эволюции, результаты отбора в эволюционном прошлом. Шансов, что такой ответ будет адекватен новым условиям, намного больше.
Чем сложнее организм, чем больше взаимосвязей проявляется в его онтогенезе, тем существенней предшествовавшая история вида, достигнутый им адаптивный компромисс будут направлять его возможные изменения. ЭТЭ позволяет нам понять, что жизнь «идёт на ощупь» (© П. Тейяр де Шарден), а не дрейфует по воле случайности.
Я благодарю за критику этого текста Александра Павловича Расницына. Я смог частично учесть его замечания, но хочу подчеркнуть, что он никак не в ответе за недостатки моих объяснений. Получилось ли у меня упомянуть всё, что важно для понимания ЭТЭ? Конечно, нет. Кое-что из того, о чем я умолчал, можно понять из презентации. Вступление (обсуждение терминов, дисклеймер) в этой колонке построено примерно так же, как в презентации, а вот в подаче самой характеристики ЭТЭ есть различия. Если хотите по-настоящему разобраться, попробуйте вникнуть и в иную версию того же объяснения.
Думать об этих вещах мне интересно. А вам?
К оглавлению
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Как в ходе эволюции возникают адаптивные признаки, или Какую из теорий эволюции подтверждают данные современной генетики? Дмитрий Шабанов
Как в ходе эволюции возникают адаптивные признаки, или Какую из теорий эволюции подтверждают данные современной генетики? Дмитрий Шабанов Опубликовано 25 ноября 2013 В последних колонках я не раз обращался к теме ЭТЭ — эпигенетической теории
Дмитрий Шабанов: Фотопериодические реакции Дмитрий Шабанов
Дмитрий Шабанов: Фотопериодические реакции Дмитрий Шабанов Опубликовано 02 ноября 2011 года Мне уже приходилось писать, что меня огорчает недооценка нашей биологической природы. Биологический фундамент, на котором вырастает и наша культура, и
Дмитрий Шабанов: Учебники. Прямиком в послезавтра Дмитрий Шабанов
Дмитрий Шабанов: Учебники. Прямиком в послезавтра Дмитрий Шабанов Опубликовано 07 апреля 2011 года Пока издательства выпускают учебники по госзаказу, доходчивость, качество, мотивирующие способности того или иного учебника будут оставаться
Дмитрий Шабанов: Блуждание глазами по всему небу Дмитрий Шабанов
Дмитрий Шабанов: Блуждание глазами по всему небу Дмитрий Шабанов Опубликовано 23 августа 2011 года Ибо весьма часто случается, что даже и нехристианин знает кое-что о земле, небе и остальных элементах видимого мира, о движении и обращении, даже
Дмитрий Шабанов: Почему есть мужчины и женщины? Дмитрий Шабанов
Дмитрий Шабанов: Почему есть мужчины и женщины? Дмитрий Шабанов Опубликовано 21 апреля 2011 года Я пообещал обсуждать биологические корни нашего поведения, а потом стал писать о другом — об учебниках, о "зелёных"... Исправляюсь.Мы так привыкли, что
Дмитрий Шабанов: Об извращённой любви к природе Дмитрий Шабанов
Дмитрий Шабанов: Об извращённой любви к природе Дмитрий Шабанов Опубликовано 18 мая 2011 года Я писал уже, что из-за активистов-природоохранников мне бывает стыдно называть себя экологом. Если вы составили представление об экологии по материалам
Дмитрий Шабанов: Энергия утреннего кофе Дмитрий Шабанов
Дмитрий Шабанов: Энергия утреннего кофе Дмитрий Шабанов Опубликовано 06 июля 2011 года Многие мои колонки посвящены биологической основе поведения людей. Увы, раз за разом я спотыкаюсь об одно непонимание. Многие читатели думают, что раз я говорю о
Дмитрий Шабанов: Цепочка следов антилопы Дмитрий Шабанов
Дмитрий Шабанов: Цепочка следов антилопы Дмитрий Шабанов Опубликовано 21 декабря 2011 года Я хочу начать с того, что некоторые задачи, которые могли бы показаться простыми, решаются чрезвычайно сложно или не решаются вообще. Для примера возьмём
Дмитрий Шабанов: Судьба точек роста в системе декоративного образования Дмитрий Шабанов
Дмитрий Шабанов: Судьба точек роста в системе декоративного образования Дмитрий Шабанов Опубликовано 05 июля 2012 года Ф-ф-ф-у-у-у-х! Учебная практика на биостанции, где я провёл последний месяц, закончилась. Прощальный момент был таков: начальник
Дмитрий Шабанов: «Экологические задачи» для студентов и школьников Дмитрий Шабанов
Дмитрий Шабанов: «Экологические задачи» для студентов и школьников Дмитрий Шабанов Опубликовано 07 марта 2012 года Учительница задаёт школьникам задачу: - У меня 36-й размер обуви, живу я на 7-м этаже и езжу на работу в школу на 42-м троллейбусе.
Дмитрий Шабанов: Парадокс ускорения эволюции Дмитрий Шабанов
Дмитрий Шабанов: Парадокс ускорения эволюции Дмитрий Шабанов Опубликовано 14 февраля 2012 года sub { font-size: 8pt; } u { text-decoration: none; border-bottom: 1px solid black; }В позапрошлый раз я обосновывал, что в движении к III синтезу эволюционная биология должна преодолеть
Первые, вторые и третьи репликаторы по Сьюзен Блэкмор, происхождение жизни и общая схема фазовых переходов в эволюции Дмитрий Шабанов
Первые, вторые и третьи репликаторы по Сьюзен Блэкмор, происхождение жизни и общая схема фазовых переходов в эволюции Дмитрий Шабанов Опубликовано 07 марта 2013 Наэлектризовавшую читателей тему двух предыдущих колонок не затрагиваю; поговорю здесь
Дмитрий Шабанов: Невидимая Нога и национальный вопрос Дмитрий Шабанов
Дмитрий Шабанов: Невидимая Нога и национальный вопрос Дмитрий Шабанов Опубликовано 24 октября 2012 года Совпали как-то в последнее время острые дискуссии, связанные с разницей национальностей и культур, идущие и в интеллектуально и культурно близкой
Эволюция эволюции: от генетического наследования к интратехнической репликации т-мемов Дмитрий Шабанов
Эволюция эволюции: от генетического наследования к интратехнической репликации т-мемов Дмитрий Шабанов Опубликовано 30 марта 2013<p Человек – не статический центр мира, как он долго полагал, а ось и вершина эволюции, что много прекраснее. Пьер Тейяр де ШарденВ этой
Краткое изложение канадских принципов
Краткое изложение канадских принципов Модельный «Свод правил по защите персональной информации», выпущенный Канадской ассоциацией стандартов, базируется на десяти независимых принципах.[260]1. Подотчетность. Организация несет ответственность за находящуюся в ее